LncRNAs play important roles in a wide range of biological processes, especially in plant reproductive development and response to stresses [40]. However, little is known about their roles in LP stress, which is a major limiting factor for plant growth and agricultural industry. Here, we undertook a genome wide ananlysis of lncRNAs in two contrasting soybean genotypes subjected to phosphate starvation.
The number of lncRNAs varies greatly across plant species. For example, 48,345 lncRNAs were identified in maize transcriptome [15], 1,212 novel lncRNAs were found in Arabidopsis seedlings grown under P-sufficient and P-deficient conditions [14]. In this study, 4,166 lncRNAs were identified. The lncRNAs identified here share most of the common features of lncRNAs reported with those in other plants, such as short length, single exons, low GC percent, which may be responsible for the common and ancient evolutionary origin. In addition, we found that the sequence length of several (70) lncRNAs were longer than 3,000 bp in soybean roots, which is similar to previous research in which 285 lncRNAs length were longer than 3,000 bp and the length of 28 lncRNAs were more than 10,000 bp [41], indicating that a small number of long lncRNAs exist in plants. The type of lncRNAs were also highly variable in plants. We identified more intergenic lncRNAs (2,865, 68.77%) than other types including antisense lncRNAs (498, 11.95%), in contrast, the number of antisense lncRNAs were greater than other types in Arabidopsis [14]. We are curious whether the number of exons is related to the length of the gene in lncRNA. After inspection, we found that the length of lncRNAs with a large number of exons did not increase significantly.
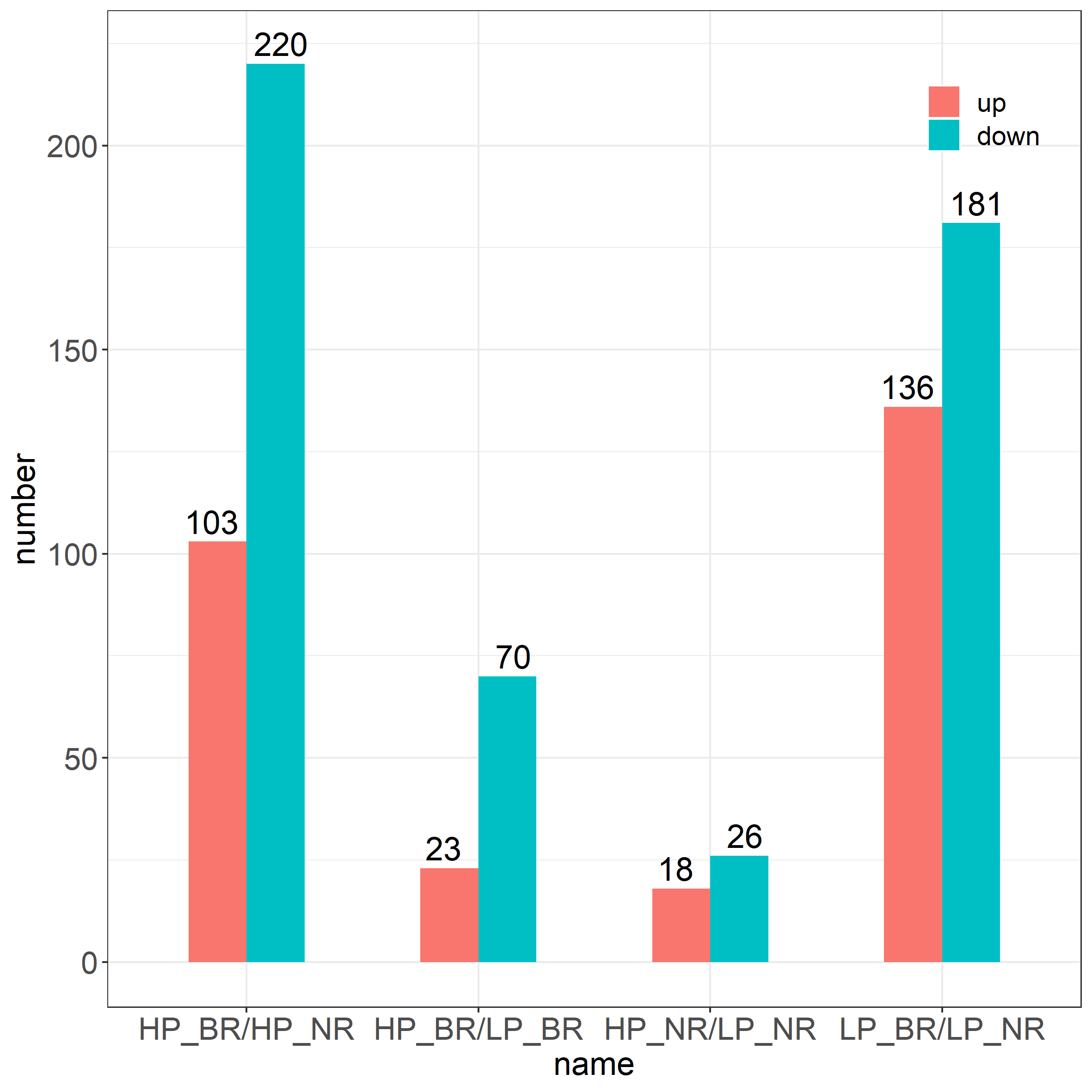
To identify the lncRNAs that were responsive to P stress, we identified differentially expressed (DE) transcripts of the lncRNAs in the pairwise comparison between two soybean genotypes under HP and LP conditions, and 525 DE lncRNAs were identified among two different genotypes in HP and LP conditions in total. As shown in Fig. 2a, Fig. 2b and Fig. S1, the number of DE lncRNAs under LP stress in Bogao is more than that in NN94156, indicating that the Bogao is more sensitive to LP treatment, which is consistent with our previous research in which Bogao was a P-sensitive genotype, while NN94156 was a P-tolerant genotype [30, 32]. Furthermore, previous and our results reveals that all the mRNA, circular RNA and lncRNA, Bogao is more sensitive to LP stress than that of NN94156 [30, 32]. The two genotypes differed greatly in response to LP stress, NN94156 and Bogao shared 21 common DE lncRNAs in HP/LP comparisons, and Bogao contained more genotype specific DE lncRNAs (72) than that of NN94156 (23) (Fig. 3b). Most of the common DE lncRNAs were consistitutively down regulated in two genotypes, indicating conserved biological mechanism for lncRNAs that involved in basal responsiveness to LP stress in both soybean genotypes. Although the similar trend of response to LP stress for these shared lncRNAs, the expression change degree of response is significantly different (Fig. 3d, Fig. 3e). To determine whether the effect of LP stress on lncRNA is related to genotypes, we compared the changes of DE lncRNAs between Bogao and NN94156 under LP or HP levels. We identified 317 DE lncRNAs (181 down-regulated, 136 up-regulated) in LP condition, suggesting that they were constitutively but differentially expressed between the two genotypes under LP condition. The 133 DE lncRNAs unique to LP condition may be play a role in LP tolerance. In contrast, 139 lncRNAs were differentially expressed in two genotypes only in HP condition, suggesting that they are differentially expressed specific to LP stress.
Studies have shown that lncRNA can directly bind to mRNA by which affecting translation, shearing, and degradation of mRNA, and can also indirectly influence the expression of mRNA [17]. Thus far, the mechanism of interaction between lncRNA and mRNA has not yet become clear. To reveal potential functions of the differentially expressed lncRNAs under LP stress in two contrast genotypes, we predicted the candidate targets of DE lncRNA, then analyzed GO terms and KEGG pathways for their putative target genes (Table S5). For DE lncRNAs in one genotype at different P levels and DE lncRNAs in Bogao or NN94156 supplied with the same P level, 403 and 1,086 GO terms with 42 and 74 KEGG pathways were significantly enriched (P < 0.05), respectively (Fig. 5). Our enrichment results showed that LP stress is a complex regulatory network involved in diverse biological processes such as lipid metabolic process, catalytic activity, cell membrane formation, signal transduction, nitrogen fixation (Fig. 5, Fig. 6), which was supported by previous studies focusing LP in soybean [32, 42]. Previous research showed that NtMYB12 acts as a phosphorus starvation response enhancement factor and regulates NtCHS and NtPT2 expression, which results in increased flavonol and P accumulation and enhances tolerance to LP stress [43]. In this study, targets of lncRNAs were enriched as KEGG pathway including flavonoid, isoflavonoid and phenylpropanoid biosynthesis, indicating that those lncRNAs may be involved in secondary metabolites to regulate P responsive genes, which needs further research.
LncRNAs can be spliced by miRNA into multiple small RNAs by which function of lncRNA can be regulated by miRNA via post-transcriptional regulation [36]. Targets of ten P related miRNAs (miR399, miR827, miR395, miR319, miR156, miR159, miR166, miR169, miR398 and miR447) were predicted, and nine of them were targets of lncRNAs as well (Table S6). Two lncRNAs, TCONS_00030280 and TCONS_00068008, possessed shared mRNA targets (Glyma.06G290000 and Glyma.12G117000, annotated as ethylene-responsive transcription factor 9-like) with the P-related miR169l-3p (Fig. 7). This finding was further supported by our recent study where NN94156 has ability to tolerate LP stress by ethylene regulator-mediated enhanced P uptake and use efficiency in roots [44]. Therefore, lncRNAs might, in part, involved in ethylene-mediated LP stress tolerance and both genes were candidate genes that merit further investigation to gain further understanding of how lncRNA involved the LP stress tolerance. Glyma.19G193900, which was predicted as purple acid phosphatase 22-like, is the target of TCONS_00105416 and miR398-x, which belonged to miR398 with demonstrated role in coping with P starvation stress [36]. Purple acid phosphatases is widely recognized as an adaption of plants to phosphorus deficiency and the secretion of PAPs (purple acid phosphatases) play important roles in P acquisition [45]. Identification of other genes with our preliminary scenario suggested that lncRNAs involve in LP stress responsive through the manipulation of genes with a variety of functionality and many of them that might also be the co-target of P associated miRNA. Detailed investigation on those genes may gain increased understanding.
Enhanced root hair production, which increases the root surface area for nutrient uptake, is a typical adaptive response of plants to phosphate starvation [46]. And ethylene plays an important role in root hair development induced by P starvation via controlling root hair elongation [37]. According to GO, KEGG and functional annotation of targets genes of DE lncRNAs, transcription factors, P-related and planthormone targets were selected to construct lncRNA-mRNA network (Fig. 8). Interestingly, we found 8 lncRNAs might be involved in ethylene regulation, as their targets were annotated as ethylene responsive transcription factor. Ethylene-responsive transcription factor belongs to APETALA2 (AP2)/ Ethylene Response Factor (ERF) family, which existed widely in plants. In Arabidopsis, the AP2/ERF TF superfamily comprises 147 members, and AP2/ERF proteins are known to regulate the responses of plants to various biotic and abiotic stresses and developmental processes. RNA interference and overexpressing of AtERF070 (AT1G71130), an ethylene response factor, resulted in altered morphophysiological traits of roots, and express change of a number of P starvation responsive genes, suggesting a potential role of this TF in maintaining P homeostasis [39]. Besides ethylene, auxin, GA and salicylic acid may be involved in response to LP stress in soybean. Induction and secretion of acid phosphatases (APases) is considered to be an important strategy for improving plant growth under conditions of low inorganic phosphate. PAPs, are an important class of plant APases that could be secreted into the rhizosphere to utilize organic phosphorus for plant growth and development [47]. Among the P-related genes, Glyma.17G172700 and Glyma.19G193900 were annotated as purple acid phosphatase. And Glyma.20G021600 were predicted as phosphate transporter, which were known as PHTs, and have important roles in P acquisition, allocation, and signal transduction. We speculate their corresponding lncRNAs may have similar functions and be involved in LP stress.
{kind=link}