Identification of the floR variants
To investigate the prevalence of the floR gene among clinical P. aeruginosa isolates, bacterial genomic DNA was pooled according to the isolated periods as TL0809 (200 P. aeruginosa strains isolated from 2008–2009) and TL151617 (230 P. aeruginosa strains isolated from 2015–2017). The Illumina sequencing of TL0809 and TL151617 generated 1.4 × 108 and 4.6 × 108 filtered reads with average depths of ~ 20 and ~ 50, respectively. The reads of TL0809 and TL151617 were assembled to generate 20,205 and 41,932 contigs, with N50 values of 3,550 and 2,316 bp in length, respectively. Using the floR gene
(AF231986) as a reference, three types of floR variants (the reference floR, floR-T1 and floR-T2, named in this study) were identified in the two pooled sequencing libraries, of which TL0809 contained all three and TL151617 contained only two floR variants (the reference floR and floR-T1) (Table 1).
Table 1.
The Abundance and PCR positive rate of the floR variants
| |
floR variants
|
Identity
|
Match length (aa)
|
Abundanceb
|
PCR positive rate
|
|
TL050607
|
floRa
|
99.3%
|
404
|
0.68
|
2/200 (1.00%)
|
| |
floR-T1
|
91.3%
|
404
|
1.18
|
6/200 (3.00%)
|
| |
floR-T2
|
87.6%
|
404
|
0.32
|
1/200 (0.50%)
|
|
TL151617
|
floR
|
99.3%
|
404
|
0.56
|
3/230 (1.30%)
|
| |
floR-T1
|
91.3%
|
404
|
4.95
|
17/230 (7.39%)
|
a reference floR (AF231986)
bAbundance = average fold of gene * 4 * 100 / average fold of 16S rRNA
Using homology tree analysis, the amino acid identities of FloR-T1 and FloR-T2 with the known floR proteins ranged from 90.80–100% and 86.10–88.90%, respectively (Fig. S1). FloR-T1 was identical to the FloR protein (YP_001715371.1) identified in Acinetobacter baumannii, while FloR-T2 showed the highest identity (88.90%) with the FloR protein (YP_005351917.1) identified in Klebsiella pneumoniae. FloRv was the floR variant with the lowest identity (88.40% − 91.80%) to other FloR proteins reported so far. However, FloR-T2 exhibited less identity with other FloR proteins than FloRv. Furthermore, FloR-T2 was shown to be one of the most divergent members of the FloR family, followed by FloRv (Fig. S1). Therefore, floR-T2 is a novel floR variant identified in this study.
The result of PCR amplification of the reference floR, floR-T1 and floR-T2 genes showed that the most prevalent variant was floR-T1, while floR-T2 was only identified in the isolates collected from 2008–2009 (Table 1). The positive rates were consistent with the abundance of the corresponding genes in the pooled genomic DNA sequencing libraries. The positive rate of the floR-T1 gene in the strains collected from 2015–2017 (7.39%, 17/230) was increased compared with that from 2008–2009 (3.00%, 6/200), which was also similar to the positive rate of the floR gene (7.01%, 23/328) of the human clinical K. pneumoniae isolates collected from the same district [5]. Zhan et al. also reported a dramatic increase in florfenicol-resistant invasive nontyphoidal Salmonella (iNTS) in China between 2007 and 2016 [3]. The results indicate that antimicrobial resistance could be transmitted from animal bacteria to human pathogens, which might pose a severe threat to public health.
Antimicrobial susceptibility of P. aeruginosa TL1285 and the floR clone
Among all P. aeruginosa strains, only one strain named TL1285, isolated from a sputum sample in 2008, carried both floR-T1 and floR-T2. P. aeruginosa TL1285 was resistant to both chloramphenicol and florfenicol (Table 2). Cloned floR-T1 (pUCP24-floR-T1/PAO1) increased the MICs for chloramphenicol and florfenicol by 2- and 4-fold, and the MICs of chloramphenicol and florfenicol against the transformant carrying floR-T2 (pUCP24-floR-T2/PAO1) increased 4- and 8-fold, respectively. The results indicate that floR-T1 and floR-T2 of P. aeruginosa TL1285 are functionally active.
Table 2
Antimicrobial susceptibility of P. aeruginosa TL1285
|
Antibiotics
|
MIC (µg/mL)
|
|
Ampicillin
|
1024
|
|
Ceftazidime
|
< 1
|
|
Levofloxacin
|
< 0.5
|
|
Cefpyridine
|
4
|
|
Minocycline
|
64
|
|
Chloramphenicol
|
128
|
|
Florfenicol
|
256
|
|
Ciprofloxacin
|
2
|
|
Azithromycin
|
32
|
|
Fosfomycin
|
256
|
|
Tigecycline
|
4
|
|
Colistin
|
< 1
|
|
Erythromycin
|
256
|
|
Nalidixic acid
|
> 1024
|
|
Gentamicin
|
> 1024
|
|
Kanamycin
|
64
|
|
Streptomycin
|
> 1024
|
Expression of the floR variants
Using transcriptome sequencing, Lang et al. found that the expression of the floR gene of the E. coli plasmid pAR060302 increased 8-fold under the induction of florfenicol [30]. Yinghui et al also reported that the mRNA levels of the floR gene encoded by ICEApl2 on chromosomes increased in the presence of chloramphenicol [31]. In this work, we performed qRT-PCR to explore the expression of the two floR variants with or without florfenicol and chloramphenicol induction. The mRNA levels of floR-T2 in P. aeruginosa TL1285 increased 78-fold and 56-fold in the presence of florfenicol and chloramphenicol, while the mRNA levels of floR-T1 only increased 1.2-fold and 1.6-fold in the presence of florfenicol and chloramphenicol, respectively (Fig. 1).
It is known that the expression of chloramphenicol resistance genes, including catA, cmlA and fexA, could be induced by chloramphenicol, and this induction is mediated by a translational attenuator at the post-transcriptional level [8, 13, 32]. The translational attenuators that located immediately upstream of these chloramphenicol resistance genes consist of a single pair of inverted repeated sequences, IR1 and IR2, as well as a reading frame of a 6–9 aa short peptide [13]. IR1 and IR2 were able to form a stable stem-loop structure blocking the resistance gene-associated ribosome binding site (RBS). Yinghui et al reported that the region upstream of the floR genes also contains a well-conserved similar attenuator sequence [31]. However, the stable stem-loop structure of the attenuator sequence did not overlap with the RBS site of the floR gene. Interestingly, we found that the peptide sequence of the floR-associated attenuator sequence in pA060302 and ICEApl2 reported by Yinghui [31] was identical to that of floR-T1 but different from that of floRv or floR-T2 by 3 amino acids (Fig. 2). Furthermore, the attenuator sequences of floR-T2 and floRv encode an identical peptide sequence, although their nucleotide sequence varies by one base. The inverted repeats of the attenuators of these floR genes were also different, and the stem-loop structures formed in distinct stable states. The genes floR-T2 and floRv have the most stable structure among the four variants. Considering that the stem-loop structure is distant to the RBS site of the floR gene, it is not clear whether this structure participates in the induced expression of the floR gene.
floR-T2 encoded in a Tn4371-like ICE
Whole genome sequencing (WGS) of P. aeruginosa TL1285 only produced a circular 6,609,407 bp chromosome with an average GC content of 66.06% encoding 5,611 ORFs. Multiple antimicrobial resistance genes, including resistance genes for β-lactams (blaOXA−50 and blaPDC−3), aminoglycosides (aadA5 and aac(3)-IIa), sulfonamides (sul1), tetracycline (tetG), chloramphenicol (catB7, floR-T1 and floR-T2) and fosfomycin (fosA), were identified in the P. aeruginosa TL1285 genome. The antimicrobial resistance phenotype was in accordance with the genotype (Table 2). The florfenicol-resistant genes floR-T1 and floR-T2 were embedded in an 86-kb Tn4371-like integrative and conjugative element (ICE) (Fig. 3). Tn4371 is a 55-kb ICE that can be integrated into the attB site (5’-TTTTCAT-3’) through a site-specific recombination process since the ends of the element can be detected covalently as a transfer intermediate [33, 34].
To track the epidemiological correlation between floR-T2 and genome islands, a BLASTN search was performed against the GenBank database using floR-T2 as a query. A total of five P. aeruginosa chromosomes, WPB099 (CP031878), WPB100 (CP031877), WPB101 (CP031876), PASGNDM345 (CP020703) and PASGNDM699 (CP020704), and one E. cloacae chromosome, AR_038 (CP030347), were found to carry floR-T2. MLST analysis of the five P. aeruginosa chromosomes together with P. aeruginosa TL1285 showed that WPB099, WPB100, WPB101, PASGNDM345 and PASGNDM699 belonged to ST308, while TL1285 belonged to ST316. P. aeruginosa ST308 is a high-risk clone that can locally acquire resistance determinants originated from the water-distribution system and caused a five-year outbreak in patients in a French hospital [35]. Interestingly, these floR-T2-carrying strains came from different resources. P. aeruginosa WPB099, WPB100 and WPB101 were isolated from hospital wastewaters in Singapore. P. aeruginosa PASGNDM345 and PASGNDM699 were derived from patient sputum in Singapore, while E. cloacae AR_038 and TL1285 were isolated from patient sputum in the United States and China, respectively. Whole genome alignment of six P. aeruginosa strains suggested that the strains shared high identity, and their differences were mainly focused on some genomic islands (Fig. 3). The Tn4371-like ICE carrying floR-T2 in TL1285 was also partially present in these five P. aeruginosa strains. It may be noted that WPB099, WPB100 and WPB101 were not fully sequenced, and the floR-T2 gene was located on an approximately 10 kb separate segment, so the precise genetic environments around floR-T2 could not be described.
Tn4371-like ICEs are mosaic in structure and consist of Ti-RP4-like transfer systems, an integrase region, plasmid maintenance genes and accessory genes [34]. Comparative analysis of the Tn4371-like ICE regions of six P. aeruginosa strains revealed that the plasmid maintenance system (repA, parA and parB) and conjugational transfer systems were conserved among these sequences (Fig. 4). The variable region between the traF and traR genes, which encoded a biphenyl catabolic bph gene cluster in Tn4371 (AJ536756), was different in these six P. aeruginosa isolates. The variable regions of WPB099, WPB100 and WPB101 were a 20-kb fragment encoding the oqxB32 gene, which confers resistance to quinolone. The variable regions of PASGNDM345 and PASGNDM699 shared high identity with those of WPB099, WPB100 and WPB101. The only difference was that in PASGNDM345 and PASGNDM699, a 13.7-kb fragment was inserted between czcD and lysR, which encode blaNDM−1, msr(E) and floR-T2 genes. The insertion fragment was flanked by 695 bp direct repeats, which were also present in WPB099, WPB100 and WPB101, whereas it showed no similarity to the existing IS element [36]. The variable region of TL1285 was similar to those of PASGNDM345 and PASGNDM699, except that the blaNDM−1-hp-msr(E) genes of PASGNDM345 and PASGNDM699 were replaced by floR-T1-tetR-tetA-lysR in TL1285. The results indicate that Tn4371-like ICEs might have emerged as a potential vehicle to mediate the spread of drug resistance genes in P. aeruginosa isolates.
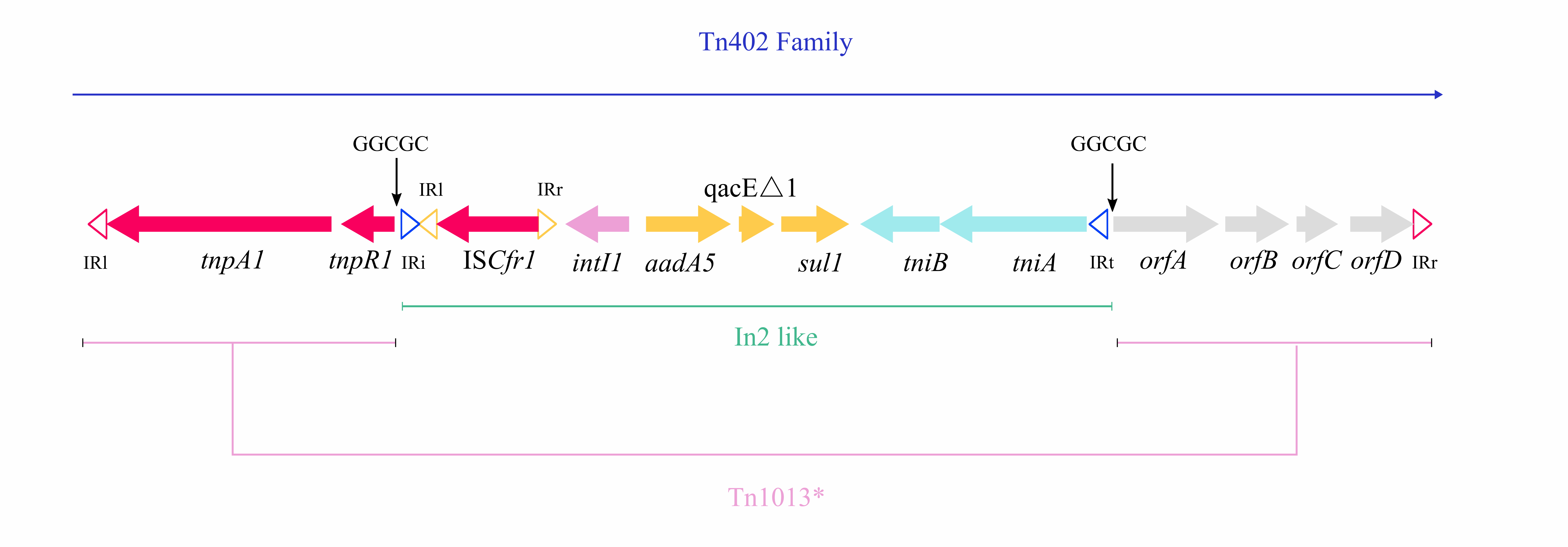
Any ICE that encodes an integrase gene closely related to intTn4371 (> 70% protein homology) and has similar maintenance and transfer genes could be considered a member of the Tn4371 family [37]. The integrase genes (int) of PASGNDM345, PASGNDM699, WPB99, WPB100 and WPB101 were identical and shared 78% identity with that of Tn4371. However, no homologue of intTn4371 was found in TL1285 (Fig. 4). Tn4371 family ICEs could be integrated into the genome through an 8-bp attB site, generating direct repeat attL and attR element chromosomal junctions [33]. In PASGNDM345, PASGNDM699, WPB99, WPB100 and WPB101, 8-bp repeats (5′-TTTTTTGT-3′) were identified on both extremities of the ICE region. However, in TL1285, only attR was found (Fig. 4). The noc gene upstream of repA in TL1285 was truncated by a novel Tn402 family transposon. The transposon is formed by ISCfr1 and In2 carrying a single aadA5 cassette embedded downstream of the tnpR gene of Tn1013, and this Tn402 family transposon was surrounded by 37-bp imperfect inverted repeats (Fig. S2).
Inverse PCR using primers P1, P2 and P3 (Fig. 4) was performed to detect whether the ICE in TL1285 could generate a circular extrachromosomal form, but no positive results were observed. Taken together, we speculate that the ICE in TL1285 is an incomplete member of the Tn4371 family and may have lost the ability to excise and integrate. The insertion of the Tn402 family transposon leads to the loss of the upstream sequence of the nor gene, including the integrase gene int of ICE.
{kind=link}