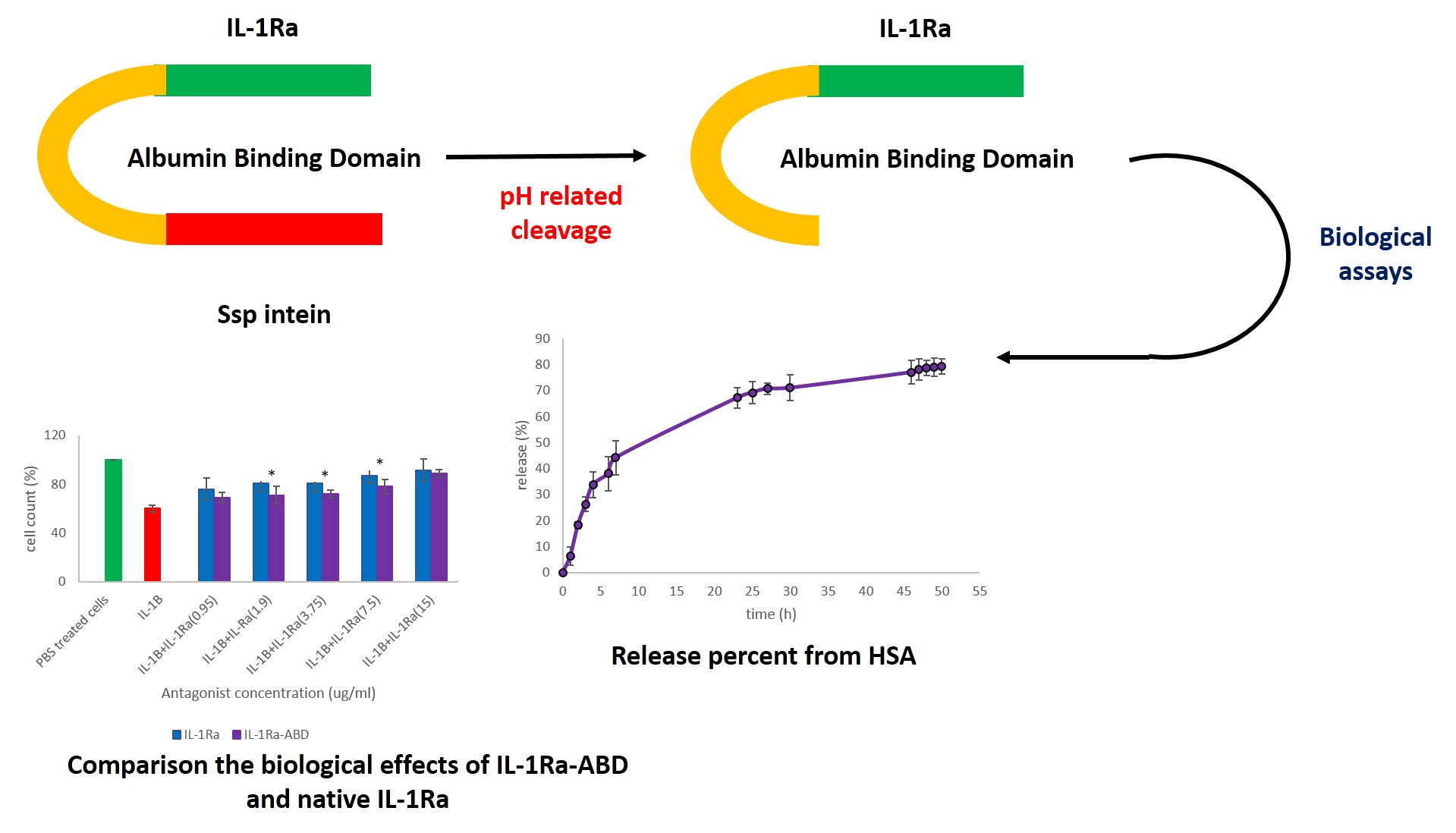
In the present study, IL-1Ra in fusing to ABD was produced in E. coli expression system in order to extend the half-life of this FDA approved drug for the treatment of RA and purified using intein mediated procedure. Our results indicated that after 2 hours, about 78 % of this fusion protein can bind to HSA and released during 50 hours until it reach to a stationary phase. On the other hand, there was not significant differences between antagonistic efficacy of IL-1Ra-ABD and native IL-1Ra used as the positive control in this test when A375 and HEK293 cell lines treated by IL-1β.
Especially during the recent years, the IMPACT system usage was increased more than ever because of the intein tags ability to produce the soluble form of recombinant proteins as well as their self-cleavage ability after the protein purification. However, optimization of the conditions for soluble expression and intein cleavage is a critical role to earn the most active protein.
Lower inducer concentrations, leads to a significant reduction in the expression of recombinant protein. For example, in one study, the effect of increasing concentrations of IPTG on the expression of DT386-BR2 protein in E. coli was evaluated. The results showed that with 1 mM IPTG, the highest amount of recombinant protein production occurs but as inclusion body [13]. In the current study, to produce the soluble protein, IPTG was used in 0.1 mM concentration according to the IMPACT manual.
As another effective variable, post-induction temperature, the formation of inclusion bodies decreases in lower temperatures [14]. For example, the human G-CSF was produced in soluble form by incubating at 15°C [15].
Although, inteins usage can significantly lower the costs of producing recombinant proteins, there are several un-solved problems. The conventional intein system, raises costs due to the need for reducing agents such as dithiotreitol (DTT) [16]. Using intein1 (ssp DnaB) of the pTWIN1 plasmid, is more convenient due to induce its self-splicing activity by changing of pH, same as the previous study for purifying the recombinant human G-CSF [15]. However, another problem in this regard is the self-cleavage induction of intein1 during the recombinant production in the host cell cytosol. In order to prevent this un-wanted reaction, lowering the post-induction temperature and the pH of cleavage buffer was used in this study. In general, cleavage ability can improve by increasing the temperature incubation of the chitin column. For example, in a study conducted by Tong, the best temperature was introduced as 37°C among various investigated temperatures range from 15 to 37°C [16]. also, there are several studies confirmed that lower pH to even 4.5, increasing the time of column incubation as well as the incubation temperature, leads to the efficient cleavage of intein1 [17]. for example, it was established in one study for the cleavage of a mutated form of insulin from the intein1 that pH 4.5 in comparison to the pH 8, and temperature as 25°C was more efficient in cleavage ratios. In our study, we used 25°C incubation temperature for 24 hours at pH 6.5 according to the IMPACT manual.
In one study which IL-1Ra was fused to human serum albumin, pharmacokinetic assays showed that native IL-1Ra was completely cleared from the blood circulation after 8 hours of injection to healthy mice, while for the fusion protein (IL-1Ra-ALB), their data represented about 30 % of the radio-labeled fusion protein could be still observed in the circulation after 48 hours after the injection. In fact, the calculated half-life of the mentioned form of IL-1Ra was determined as about 10 hours in comparison to the native form with calculated half-life as about 0.34 hours [9]. In our study, we used ABD instead of full-length human serum to overcome the probable drawbacks of expression the protein in fusion to a large molecule.
ABD usage for extending the half-life of biological drugs was used in several studies mentioned in introduction section. For example, in attempt to increase the half-life of exenatide using ABD, the pharmacokinetics assays showed about 32-fold increasing in the half-life of exenatide-ABD when compared to native form (16 hours vs. 30 min) [10]. Our in vitro release test, confirmed that IL-1Ra fused to ABD could release from the human serum albumin until 50 hours and Native-PAGE revealed the stability of this fusion protein.
For biological assay, on the other hand, we used the potent cytotoxic and apoptotic effects of IL-1β against cells with highly expressed of IL-1R [19]. Furthermore, based on the effects of Anakinra in preventing the cytotoxic effects of IL-1β, the protocol mentioned in the methods section was used. The biological results showed more increase in the concentration of IL-1Ra-ABD, more inhibition in effects of IL-1Ra on IL-1β. In the study of Lui et al, this method was used for evaluating the inhibitory effects of a fusion protein of IL-1Ra with an extended half-life. The results showed that 32 nM of IL-1Ra led to 100 % inhibition effects of 1 ng/mL IL-1β [19]. The concentrations of IL-1Ra-ABD used in the present study were higher than those investigated by Lui et al. In another study, Yu-Xin produced several mutated forms of IL-1Ra and compared their biological activities with those of native IL-1Ra. The results showed that at higher investigated concentrations (25 µg/ml), IL-1Ra can inhibit the effects of IL-1 [20].
The antagonizing efficacy of IL-1Ra-ABD was not statistically significant compared to native IL-1Ra for highly expressed IL-1 receptor cells, A375. The strength of the produced recombinant protein was about 101% of the alone form.
Lui et al., evaluated the antagonizing efficacy of the recombinant IL-1Ra produced in Pichia pastoris and said that this protein could antagonize the cytolytic activity of IL-1β (1 ng/ml) when added only after the treatment of cells by IL-1β [21]. However, in the present study, based on our previous project on evaluation the antagonizing efficacy of IL-1Ra, produced with the same protocol, a time interval of one hour was used. The results showed that IL-1Ra-ABD could successfully act in antagonizing the toxic effects of IL-1β in a time interval of 1 hour without any significant differences to native IL-1Ra.
Finally, Powers et al. used IL-1-responsive A549 cell line for evaluating the competitive antagonizing effects of a Pasylated form of IL-1Ra, produced for expanding its half-life [21]. In this study, the antagonist was added to cells and 1 hour later, IL-1α was added to the cells. The efficacy of IL-1α (in the final concentration of 10 ng/ml), in IL-6 release was evaluated and found that both Anakinra and its Pasylated form antagonized the IL-6 secretion [22]. The results of this study, also confirmed the Anakinra antagonizing effects on IL-1, only before its internalizing to the cells with highly expressed receptors on their surface.
{kind=link}