Human Sirt2 can be SUMOylated in vitro and in vivo.
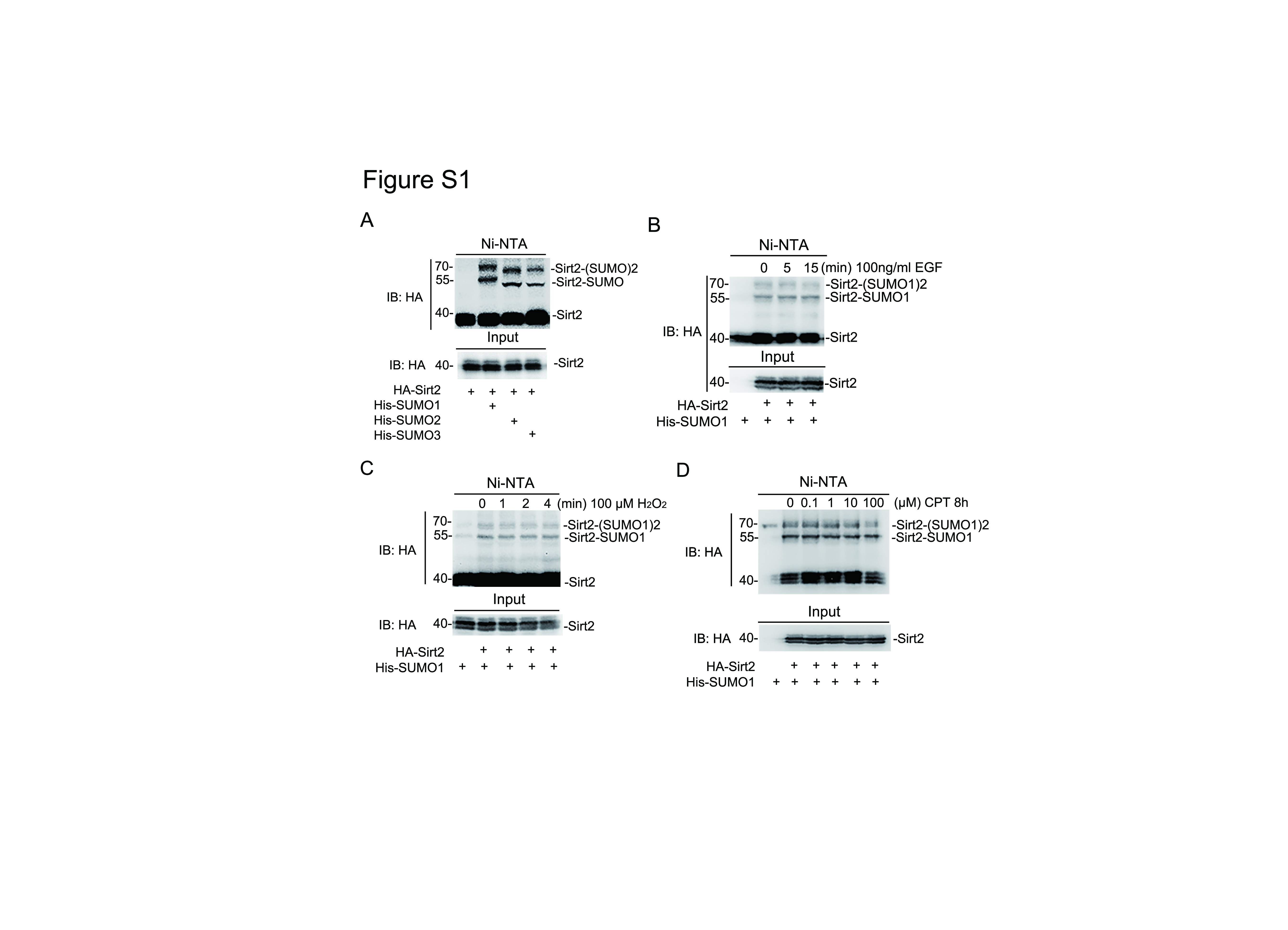
To determine whether Sirt2 can be SUMOylated, HEK293T cells were used to over-express HA-tagged Sirt2 with or without His-tagged SUMO1 and Flag-tagged UBC9, cell lysis after transfection were subjected to Ni-NTA pull-down assay and western-blot analysis. Two major shifted Sirt2 protein bands were observed and their levels could be enhanced or attenuated by E2 enzyme UBC9 or deSUMOylation enzyme SENP1, respectively (Fig. 1A). In consistent with that, the immunoprecipitation assay also showed a same outcome about Sirt2 SUMOylation events in HEK293T cells (Fig. 1B). To identify the endogenous SUMOylation on Sirt2 protein, SH-SY5Y cells stably expressing HA-Sirt2 were harvested and subjected to immunoprecipitation with HA antibody, which showed that Sirt2 SUMOylation indeed could happen in vivo according to the detection by SUMO1 antibody and HA antibody (Fig. 1C). In the prokaryotic expression system, in vitro GST-pull down assay also proved that GST-tagged SIRT2 could be SUMOylated when co-transfected with SUMO1, E1 and E2 enzymes in BL21 E. Coli host (Fig. 1D). The endogenous SUMOylation on Sirt2 was also identified in SH-SY5Y cells through immunoprecipitation assay with SUMO1 and Sirt2 antibodies (Fig. 1E). In addition to SUMO1 modification, SUMO2 and SUMO3 modifications were also positively identified in HEK293T cells (Fig S1A).
As an NAD+-dependent deacetylase, we wonder whether Sirt2 SUMOylation could be stimulated by NAD+ in cells. Surprisingly, 1 mM NAD+ could obviously enhance the Sirt2 SUMOylation level but higher NAD+ concentration would not help the further improvement (Fig. 1F). However, it had a significant and positive time effect on Sirt2 SUMOylation under 1 mM NAD+ stimulation (Fig. 1G). On the contrary, EGF, H2O2, or even cisplatin (CPT) did not affect the SUMOylation level of Sirt2 (Fig S1B-D). Together, these evidences indicate that Sirt2 can be SUMOylated in vitro and in vivo, which can be specifically stimulated by NAD+.
Sirt2-SUMOylation does not affect the localization and stability of Sirt2 in cells.
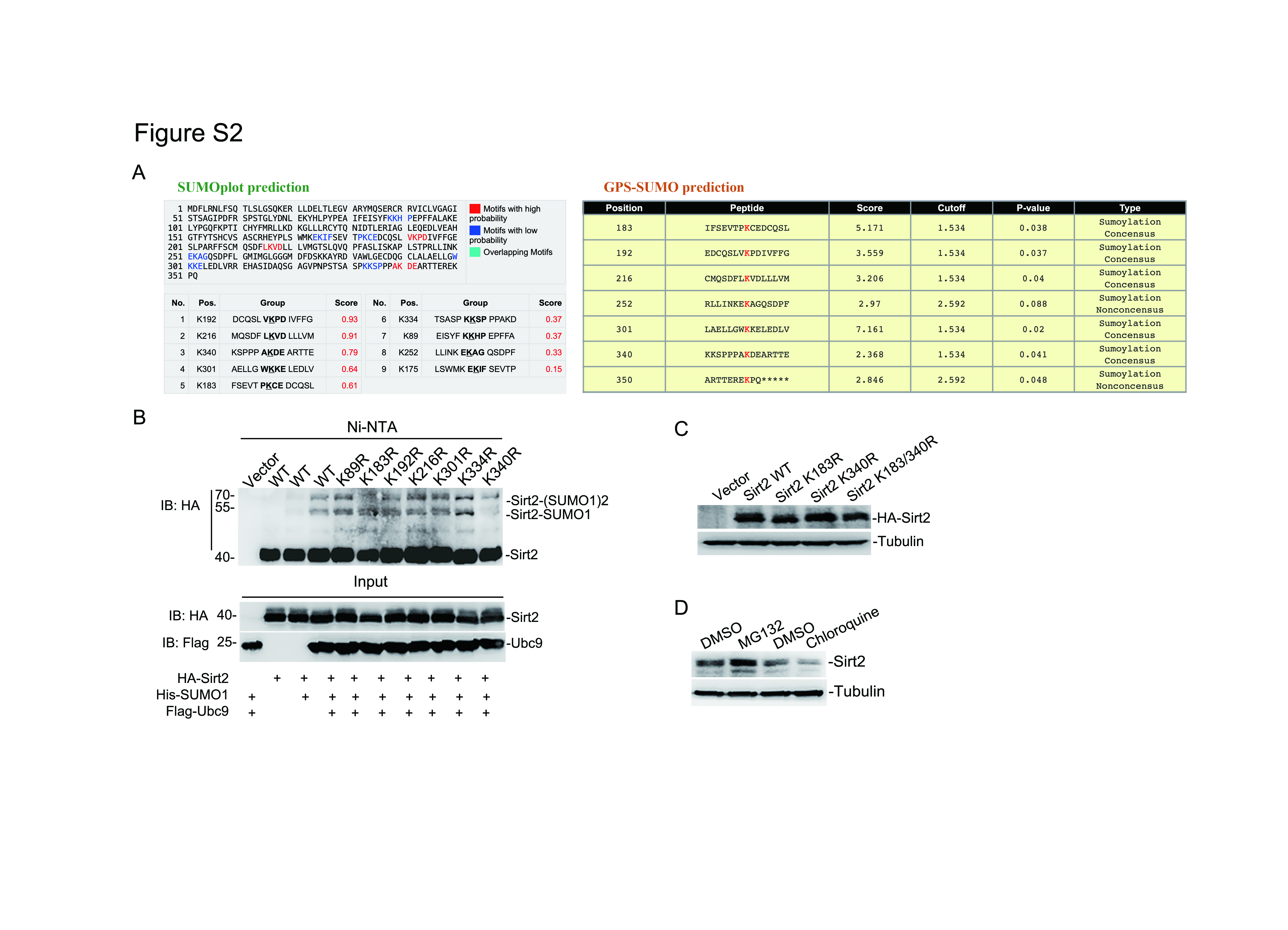
To determine which lysine was conjugated by SUMO1 molecule, SUMOplot™ Analysis Program (http://www.abgent.com.cn/tools/sumoplot) and GPS-SUMO 2.0 Online Service (http://sumosp.biocuckoo.org/online.php) were applied to predict the probability of SUMOylation sites within Sirt2 protein (Fig S2A). There were six potential SUMOylation sites predicted by both software with the different system approaches [30], including Lys183, Lys192, Lys216, Lys252, Lys301 and Lys340 (Fig. 2A and Fig S2A). Sequentially, substitution of these lysine (K) to arginine (R) respectively revealed that both K183R and K340R significantly attenuated the SUMOylation level of Sirt2 (Fig S2B and Fig. 2B). Intriguingly, the K183R mutation within the Sirt2 protein notably removed a shifted protein band but enhanced the level of another one, however, the K183R and K340R double mutations caused a significant decline of the total Sirt2 SUMOylation level according to both of the Ni-NTA pull-down and immunoprecipitation assays (Fig. 2B & C).
To assess whether SUMOylation on Sirt2 can affect its localization and stability in the cell, HA-tagged Sirt2 wide type (WT) or mutants were cloned into pGreenPuro-Dual vector (see description in Fig. 3G) and transferred into SH-SY5Y cells through lentiviral infection, respectively, and the even expression level was identified by western-blotting (Fig S2C). The nucleus/cytosol extraction assay revealed that SUMOylation did not alter the localization of Sirt2, whatever on K183, K340, or K183 + 340 (Fig. 2D & E). Moreover, the Immunofluorescence assay also showed a consistent outcome (Fig. 2F). Sirt2 protein stability was controlled by the proteasome system but not lysosomal pathway according to the protein level detection in cells after MG132 or Chloroquine treatment (Fig S2D), yet, the SUMOylation on Sirt2 did not significantly affect its stability by detecting the protein half-life after cell-treating with cycloheximide (CHX) (Fig. 2G). Taken together, these data indicate that both K183 and K340 are the main SUMOylation sites in Sirt2, but the SUMOylation on Sirt2 impacts neither of its localization or stability in neuroblastoma cells.
Sirt2-SUMOylation mainly inhibits P38-mTORC2-AKT signaling in neuroblastoma cells.
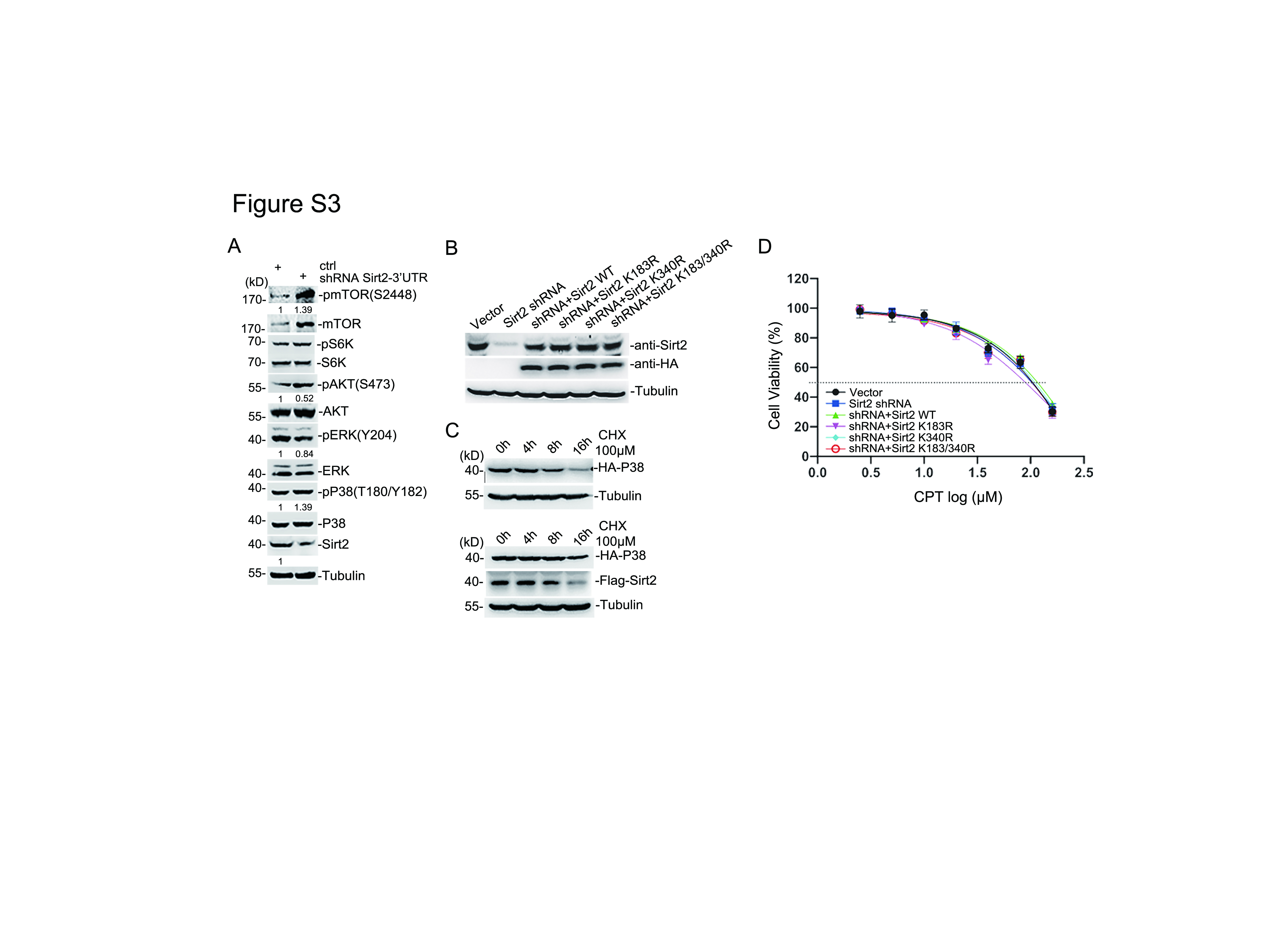
As an important deacetylation enzyme, Sirt2 participates in various cell activities such as aging, cell signal transduction and carcinogenesis [31], however, its signaling mechanisms in cells are still not well understood. Considering this, we silenced the expression of Sirt2 in SH-SY5Y cells through pGreenPuro-dual lentiviral system with the specific shRNA. The pP38, pAKT and pmTOR were significantly enhanced but pERK was short-term depressed when abrogating the Sirt2 expression after starvation following with complete culture medium stimuli (Fig. 3A left two lanes in 3B, Fig S3A). mTORC1 and mTORC2 are the two main sensors of mTOR signaling, while the activation of pS6K (pP70)/p4E-BP1 or pAKT reflects the mTORC1 or mTORC2 signaling pathway, respectively. Here, Sirt2 silencing did not significantly enhance the activation of p4E-BP1 but did only slightly impact on pS6K (Fig. 3A, supplemental Fig S3A). These data indicated that Sirt2 mainly involved in the P38, AKT and mTORC2 signal transduction in neuroblastoma cells. To clarify the regulation role between P38 and mTORC2/AKT caused by Sirt2 abrogation, several inhibitors were applied for further analysis (Fig. 3B-E). Rapamycin as mTOR specific inhibitor can inhibit mTORC1 activation but has no effect on mTORC2, and it did not affect the level of pP38, pmTOR and pAKT under the Sirt2 silencing condition (Fig. 3B middle 4, 5 lanes & C). Also, AKT inhibitor (LY294002) did not impact the P38 activation, but significantly abolished the pmTOR and pS6K level (Fig. 3B right 3,4 lanes & C). Intriguingly, both pAKT and pmTOR levels were suppressed when P38 signal was blocked by SB202190 (Fig. 3B right 1, 2 lanes & C). In addition, pp242, an inhibitor of both mTORC1 and mTORC2, was applied to confirm that mTOR signal indeed did not affect the Sirt2-associated P38 activity (Fig. 3D & E). These data indicate that Sirt2 in neuroblastoma can mainly suppress P38-mTORC2-AKT signaling (Fig. 3A-E). The strategy of cell signal transduction mediated by the specific inhibitors showed that Sirt2 can only slightly or temporarily affect the mTORC1 or ERK signaling, but is mainly responsible for suppressing the P38-mTORC2-AKT signaling in neuroblastoma (Fig. 3F).
To rescue Sirt2 expression in SH-SY5Y cells harboring silenced endogenous Sirt2 for re-observing the signal transduction, we constructed a homemade lentivector based on pGreenPuro™ shRNA lentivector (System Biosciences, Switzerland), termed pGreenPuro-Dual, which contained two independent expression cassettes generating non-coding RNA transcripts (i.e. shRNA, miRNA or lncRNA) by H1 promoter, and producing mRNA for translation by CMV promoter, respectively (Fig. 3G). This all-in-one lentiviral vector can be used for recombinant lentivirus packaging to silence endogenous gene and express exogenous gene (i.e. wide type or mutants) with only once infection force on the target cell, which avoids the uncontrollable changes of cellular behavior caused by multiple viral infections and drug selections.
Then, the SH-SY5Y cells were subjected to infection by recombinant lentivirus harboring shRNA targeting to endogenous Sirt2-3’UTR and rescued exogenous HA-Sirt2-WT, -K183R, -K340R or -K183/340R, and cells infected with virus generating by pGreenPuro-Dual vector or pGreenPuro-Dual/shRNA were set as the system control. After puromycin selection, the endo- or exo-Sirt2 protein expression level in those cell lines was quantified by western-blotting (Fig S3B). Surprisingly, re-introduced Sirt2-WT suppressed the activation of P38, mTOR and AKT resulting from endo-Sirt2 ablation, while SUMOylation-deficient Sirt2 (K183R, K340R or K183/340R) can not resume the normal signal transduction in SH-SY5Y cells (Fig. 3H & I). Taken together, these evidences indicate that Sirt2-SUMOylation is critical in inhibiting the P38-mTORC2-AKT signaling in neuroblastoma.
Sirt2 is a promised deacetylase of MAPK/P38 and its enzyme activity relates to SUMOylation.
Acetylation of lysine residues within proteins is relevant to the physiological activities of targeted proteins. P38 was proved to be reversibly acetylated by PCAF/P300 and deacetylated by HDAC3, which regulates its affinity for ATP binding and P38 kinase activity [32]. However, there have no evidence yet to illuminate the relationship between Sirt2 and P38. To investigate whether Sirt2 could directly regulate P38, the Immunoprecipitation assay revealed that Sirt2 could interact with P38 in cells (Fig. 4A). In consistent with that, the GST-pull down assay also proved that GST-Sirt2 recombinant protein could specifically bind to endogenous P38 in SH-SY5Y cells, but GST-Sirt1 protein could not (Fig. 4B). As one of the deacetylation enzymes, we wonder whether Sirt2 can regulate the acetylation level of P38. The immunoprecipitation assay showed that Sirt2 could obviously decrease both of the exogenous and endogenous P38 acetylation levels (Fig. 4C & D). Comparing with the known deacetylase HDAC3, Sirt2 showed a similar deacetylation ability to P38 protein (Fig. 4E). Moreover, the P38 acetylation level was obviously enhanced by treating with Sirt2-specific inhibitor AK-7 (Fig. 4F). Traditionally, acetylated proteins are more stable comparing to their unacetylated counterparts, because unacetylated lysines are usually targeted for ubiquitination-mediated proteasomal degradation [33]. Considering of this, we compared the half-life of P38 with or without Sirt2 expression, the result conversely showed that deacetylated P38 was more stable than the acetylated one (Fig. 4G & Fig S3C), which was consistent with a certain phenomenon that Sirt2 deficiency can stabilize NFATc2 to enhance its transcription activity [34]. To assess whether Sirt2-SUMOylation could affect its deacetylation activity, the SH-SY5Y stable cell lines harboring Sirt2-WT or -mutants were used for immunoprecipitation with acetyl-lysine antibody and the acetylated-P38 level indicated that SUMOylation significantly impaired the deacetylation ability of Sirt2 (Fig. 4H). Thus, these results together suggest that Sirt2 is a promised deacetylase of P38 and the enzyme activity is closely associated with its SUMOylation.
Sirt2-SUMOylation is critical in suppressing the phenotypes of neuroblastoma cells.
To assess the phenotypes induced by Sirt2-SUMOylation in neuroblastoma cell, we first detected the proliferation ability of SH-SY5Y cell lines with silenced Sirt2 or directly overexpressed Sirt2 variants (i.e. WT, K183R, K340R or K183/340R), which showed Sirt2 ablation or Sirt2-SUMOylation deficiency significantly facilitated the proliferation capability, but Sirt2-WT showed an opposite effect (Fig. 5A). Meanwhile, re-applied Sirt2-WT but not Sirt2-K183R, -K340R or -K183/340R resumed the proliferation in Sirt2-silenced cells (Fig. 5B). According to the soft-agar colony-forming assay, knock-down Sirt2 or overexpress Sirt2 mutants (K183R, K340R, or K183/340R) enhanced the clone formation, yet overexpressed Sirt2-WT significantly suppressed the clone formation (Fig. 5C upper panel and D). Similarly, re-applied Sirt2 variants in Sirt2-deficient cells showed almost the same outcomes (Fig. 5C lower panel and E). To inspect the cell mobility and invasiveness resulted from Sirt2-SUMOylation, the migration and invasion assays were conducted by using the cell chamber in 24-well plates. The results revealed that Sirt2 can significantly suppress the migration and invasion abilities of SH-SY5Y cells (Fig. 5F-K). But SUMOylation-deficient Sirt2, whatever overexpression (Fig. 5F, I upper panel & G, J) or rescued expression (Fig. 5F, I lower panel & H, K), intensified the cell capabilities of migration and invasion in vitro. In addition, we wonder whether Sirt2-SUMOylation affected the drug susceptibility, the IC50 of Cisplatin and AK-7 were detected in the mentioned cell lines above, respectively. The result showed that Cisplatin, as a frequently used chemical drug for neuroblastoma, did not have any different effect among SH-SY5Y cells with or without Sirt2 or its mutants comparing to control (Fig S3D). While AK-7 proved to be able to inhibit Sirt2 activity and enhance the phosphorylation of P38 in SH-SY5Y according to our previous report [35], showed a significant different drug-effect between SH-SY5Y cells harboring Sirt2-WT and those harboring Sirt2-mutants (K340 mainly) (Fig. 5L). Together, these data reveal that Sirt2-SUMOylation plays an important role in inhibiting the malignant phenotypes in neuroblastoma cells and makes a contribution to the cell sensitivity to Sirt2-specific inhibitor AK-7 in vitro.
Sirt2-SUMOylation exerts tumor-suppressor property in mice and Sirt2 expression is closely related to the patient’s prognosis.
In xenograft mouse model, the tumor size or weight was measured at 2 weeks after subcutaneous injection. Comparing with vector or mock group, both of weight and size of the tumors from the Sirt2-deficient or -mutants cell injection group were much greater. However, the overexpressed Sirt2-WT significantly suppressed the tumor growth (Fig. 6A & B). To trace expression changes of Sirt2 in the development and progression of neuroblastoma, a series of statistical analyses were performed using the R2 online database (https://hgserver1.amc.nl/cgi-bin/r2/main.cgi). Comparing to normal brain tissue, the Sirt2 expression is significantly suppressed in tumors from three different datasets (Fig. 6C). The survival curve also revealed that lower Sirt2 expression predicted a poor survival probability of either event-free or overall (Fig. 6D & E). Moreover, with the increase of tumor pathological stage, the expression of Sirt2 showed a downward trend according to the R2 dataset analysis, particularly in stage 4 (Fig. 6F). Moreover, neuroblastoma from stage 4 s which is similar to those from stage 1, 2 in molecular [36] showed the same Sirt2-expression pattern (Fig. 6F). Taken together, these all indicate that Sirt2 expression is closely associated with the development of neuroblastoma and the patient’s prognosis. As well, SUMOylation on Sirt2 seems essential for suppressing the tumorigenesis of neuroblastoma in mice.
{kind=link}
{kind=link}
{kind=link}