The paper presents genetic characteristics of two strains of the spring viraemia of carp virus strains Kirov/08 and Orenburg/14 isolated in the Kirov and Orenburg Oblasts of the Russian Federation, respectively. The nucleotide sequence analysis of the 516-bp fragment of Kirov/08 and Orenburg/14 G gene showed 9.5% difference. The phylogenetic analysis demonstrated a high level of homology of these strains to SVC viruses, isolated earlier in Russia, Ukraine and the Republic of Moldova.
Spring viraemia of carp (SVC) is an acute highly contagious viral disease of cyprinids grown in fish farms and inhabiting natural reservoirs [1]. According to the classification of the World Organization for Animal Health (OIE) SVC belongs to highly dangerous notifiable fish diseases [13].
The SVC causative agent is an RNA-containing bullet-shaped virus (SVCV) with the particle size of 105-125×70-85 nm belonging to the genus Sprivivirus in the family Rhabdoviridae of the order Mononegavirales [1, 6, 9]. The virus genome is a non-segmented, negative-sense, single-stranded RNA approximately 11 kb in length encoding five major structural proteins: N (nucleocapsid protein), P (phosphoprotein), M (matrix protein), G (glycoprotein), and L (polymerase) [10].
The virus causes disease in several carp species. Roach (Rutilus rutilus), bream (Abramis brama), tench (Tinca tinca), bluefin (Thunnus thynnus), catfish (Silurus glanis), pike (Esox lucius), white cupid (Ctenopharyngodon idella), silver carp (Hypophthalmichthys molitrix) and bighead carp (Aristichthys nobilis) can also be infected. Juveniles under one year of age are particularly susceptible to the disease, but fish of any age can become infected [3]. The disease usually occurs in spring (from April to early June), but sometimes it is recorded in autumn.
SVC is characterized by the development of a septic process manifested as an exudative-hemorrhagic syndrome as a result of virus propagation in the endothelium of blood capillaries and kidneys, which leads to disrupted water-mineral balance and the release of plasma and shaped blood elements into the surrounding tissues and body cavities [5]. The mortality rate can reach 70% depending on the age and general health of fish population. [3].
The SVCV was first identified on the territory of the former Yugoslavia by N. Fiyan in 1968 and in Russia - by N.I. Rudikov in 1971 [1] and is currently widespread in the regions of carp breeding in Europe and Russia. The virus is also reported in South and North America, China and Korea [3, 11, 14, 15]. The last outbreaks of this disease in Russia were registered in 2008 and 2014 in Privolzhsky Federal District (Kirov and Orenburg Oblasts) [1]. Data on global distribution of the SVCV are shown in Figure 1.
The spread of the SVCV results in significant economic damage, including losses due to fish mortality, costs related to implementation of animal health and quarantine measures, and trade restrictions [1]. There is no effective treatment for the disease, so prevention is the best way of control [3, 7].
In Russia the SVC diagnosis is currently made based on analysis of epidemiological data, the disease clinical signs, study of post-mortem lesions, supported by test results based on virus isolation in susceptible cell lines (EPC – epithelioma papulosum cyprini, FHM – fathead minnow or GCO – grass carp ovary). In addition, serological identification of the virus antigen, followed by a bioassay and detection of viral RNA using polymerase chain reaction (PCR) is applied.
Research studies done in recent years [2, 4, 10, 17, 18] showed that PCR used for the SVC diagnosis, in addition to virological and serological methods, significantly expands the possibilities for rapid identification of the virus. In addition, PCR followed by sequencing of amplified regions of the viral genome allows carrying out phylogenetic analysis, differentiation of the SVCV genetic variants and studying of molecular epidemiology of the disease.
Thus, the aim of this paper is to perform molecular analysis of two SVCV strains Kirov/08 and Orenburg/14 detected in the Kirov and Orenburg Oblasts, respectively, and to study their phylogenetic relations.
The strains Kirov/08 and Orenburg/14 were isolated in 2008 and 2014 from samples obtained from mirror carp caught in fishery farms of the Kirov and Orenburg Oblasts of the Russian Federation. The both strains were freeze-dried and deposited in the collection of strains of microorganisms of the FGBI "Federal Center for Animal Health" (FGBI "ARRIAH", Vladimir, Russia).
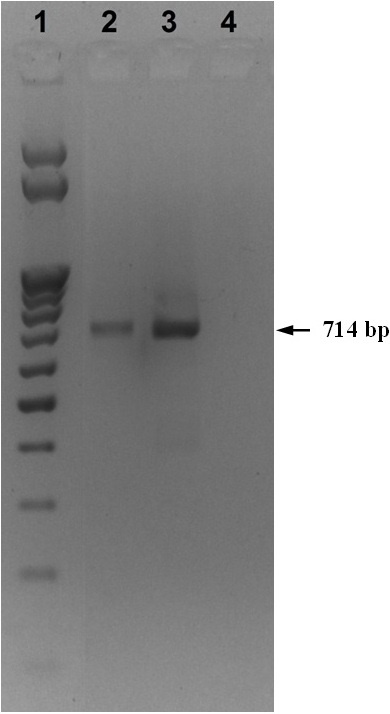
The total RNA was isolated from both SVCV strains using ExtractRNA commercial reagent (ZAO Eurogen, Russia) (a monophasic aqueous solution of phenol and guanidine isothiocyanate) according to the manufacturer's instructions. REVERTA-L commercial reagent kit (OOO InterLabService, Russia) was used according to the manufacturer's instructions to obtain cDNA on the matrix of the isolated RNA. The obtained cDNA was used for PCR, which was carried out in PCR Sprint thermal cycler (Thermo Scientific, USA) according to the generally accepted method under the following modes: denaturation- for 1 min at 95° C, annealing of primers – for 1 min at 57° C, synthesis – for 1 min at 72oC for 35 cycles. At the final stage, the mixture was heated for 10 minutes at 72° C. The reaction mixture for PCR was collected at a volume of 25 µl using Encyclo Plus PCR commercial reagent kit (ZAO Eurogen, Russia), according to the manufacturer's instructions. The following primers were used for SVCV identification: SVCV-F1 5’-TCT-TGG-AGC-CAA-ATA-GCT-CAR-RTC-3' and SVCV-R2 5'-AGA-TGG-TAT-GGA-CCC-CAA-TAC-ATH-ACN-CAY-3', previously proposed by D. M. Stone et al. [18] allowing amplification of a fragment of the G gene encoding the glycoprotein protein. The 714 bp PCR product was analyzed by 1.5% agarose gel electrophoresis. M25 (NPO SibEnzim, Russia) was used as a molecular weight marker.
Sequencing was performed using 3130 DNA Analyzer (Applied Biosystems, USA) and BigDye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems, USA) according to the guidelines for DNA sequence determination using sequence analysis with Applied Biosystems 3130 and 3130xl genetic analyzers based on automation of sample preparation of sequencing reaction products using the Freedom EVO-2 100Base unit (FGBI “ARRIAH”, Vladimir, 2012). The obtained nucleotide sequences were analyzed using the BioEdit v.7.0.5.3. software and submitted to the GeneBank database with the following accession codes: MZ531862 for Kirov/08 and MZ555809 for Orenburg/14.
The phylogenetic analysis of the nucleotide sequences obtained during the study and received from the GenBank (NCBI) international database was performed using the MEGA 6.0 Software [19] based on the maximum likelihood method. Bootstrap support was 1000. Nucleotide sequence alignment was carried out using the ClustalW algorithm. Data on the SVCV strains and isolates, including their indication and number in GenBank are presented in the Supplementary table.
The G gene-encoded glycoprotein forms trimeric peplomers, or spikes, on the surface of the SVCV, which bind to cellular receptors and cause viral endocytosis. It also carries neutralizing epitopes and is a potential target for the development of DNA vaccines. To date, glycoprotein is the most important antigen for determining the serological properties of rhabdoviruses, and the G gene is the most variable within the SVCV genome [8]. This may explain the fact that the G gene was used for phylogenetic analysis of SVCV strains and isolates by Basic A. et al. [8], Sheppard A.M. et al. [16], Stone D. M. et al. [18], Vicenova M. et al. [20] and Warg J. V. et al. [21]. In this regard, we used primers amplifying the SVCV G gene region considered as the most informative for genetic analysis.
According to the phylogenetic analysis conducted by Stone D. M. et al. [18] based on the nucleotide sequence of the G gene region, all SVCV isolates and a number of related viruses were classified into 4 genogroups. Genogroup I contains classical SVCV isolates, Genogroup II - GrCRV isolates (grass carp rhabdovirus), Genogroup III – PFRV isolates (pike fry rhabdovirus) and Genogroup IV – TenRV isolates (tench rhabdovirus). Genogroup I isolates were further divided into 4 subgroups (a, b, c and d) according to their geographical origin, and a high rate of difference in the nucleotide sequences of the G gene region. Subgroup “a” includes isolates from Asia and strains introduced to Europe and North America, subgroups “b” and “c” comprise isolates from Eastern Europe and Russia, and subgroup “d” – isolates from Western Europe.
Analysis of the nucleotide sequences of the G gene fragment 516-bp in length revealed that the Kirov/08 and Orenburg/14 SVCV strains differed at 49 positions (9.5%), which indicates significant difference between them. Only Genogroup I SVCV strains were used for phylogenetic analysis in the study.
The generated phylogenetic tree (Fig. 2) showed that all SVCV strains and isolates analyzed in the study formed four clusters earlier identified by Stone D. M. et al. [18]. The strain Kirov/08, with a high bootstrap value of 99%, was clustered with isolates from subgroup “c”: N1-5 isolated in Ukraine and P4 isolated in Russia. Orenburg/14 strain, clustered with high bootstrap values (97% and 88%) with the RHV and 2/90 strains from Ukraine and the Republic of Moldova, respectively, and belonged to subgroup “b”. Thus, the phylogenetic analysis results showed that the Kirov/08 and Orenburg/14 SVCV strains belong to the group of strains previously isolated in Eastern Europe and Russia.
The phylogenetic analysis of the studied Kirov/08 and Orenburg/14 strains revealed that both of them belong to SVCV Genogroup I and allowed determining the relatedness of the strains isolated in Eastern Europe and Russia: the Kirov/08 strain belongs to subgroup “c” and Orenburg/14 strain belongs to subgroup “b” of SVCV Genogroup I. The data obtained will allow performing of molecular-epidemiological studies in case of future outbreaks of SVC in Russia.
{kind=link}