Autoimmune thyroid disease (AITD), which encompasses two types: autoimmune hyperthyroidism known as Graves’ disease (GD) and autoimmune Hashimoto’ s thyroiditis (HT), is a frequent autoimmune disease caused by the autoimmune dysfunction, which is characterized by the infiltration of lymphocytes in the thyroid and the production of thyroid autoantibodies (1). The pathogenesis is complex, involving heredity and environmental factors. And the incidence is higher in women than in men, increasing with age. Genetic factors, radiation exposure, increased iodine intake, infection, smoking, life stress, and the use of amiodarone and interferon can lead to AITD in susceptible individuals, of which 70% can be attributed to genetic factors. The common model of the onset of AITD is the underlying genetic susceptibility and triggers that trigger and maintain the autoimmune response, which ultimately leads to hypothyroidism or hyperthyroidism. It is well known that thyroid antibody expression positive play an important role in the diagnosis of AITD, and there is growing concern about the high positive rate of thyroid autoantibodies in the population and its adverse effects on human health, including an increased risk of cardiovascular disease and death, an increased rate of abortion among women of childbearing age and an impact on the fetal mental development. However, due to its known multiile causes and slow and complicated autoimmune process, we have no good prevention and control measures for AITD.
HT manifests as subacute or destructive thyroiditis and its pathogenesis is thought to be driven by th1 cells.T cells can directly damage the thyroid gland, and the release of thyroid hormones can lead to hyperthyroidism, followed by exposure to thyroid antigens (thyroid peroxidase and thyroglobulin) leading to antibody production and ultimately hypothyroidism. Thyroid gland dysfunction may manifest as clinical thyroid dysfunction (0.1-2% of the population) or subclinical thyroid dysfunction (10–15% of the population), depending on the extent of damaged thyroid parenchyma, but HT patients may also have normal thyroid function(3). The major autoantibodies in the peripheral blood of HT patients are thyroid peroxidase antibody (TPOAb) and thyroglobulin antibody (TgAb), but they also occur in about 70% of GD patients. Thyroid peroxidaseantibody (TPOAb), which indicates thyroid lymphocyte infiltration and follicular epithelial destruction, is detectable in 90% of HT patients and in 5–24% of the general population(2)(4). Nearly 50% of patients diagnosed with HT due to TPOAb positivity have normal thyroid function, while the majority of other patients have subclinical (mild) hypothyroidism, which is characterized by normal FT4 levels and elevated serum TSH, and only a few patients have severe hypothyroidism. It is estimated that about 2.5% of people with TPOAb-positive can develop noticeable hypothyroidism and about 4.5% of people with TPOAb-positive and elevated TSH can develop significant hypothyroidism annually(5). TPOAb status (positive or negative) is important in the initial diagnosis of patients, and both the presence and concentration of TPOAb are associated with the beginning of the disease. One study showed that TPOAb increased the adverse risk at > 500kU/l, while there was no significant difference in the adverse risk at its low levels(6). The exact function of TgAb is unclear, as they do not cause thyroid cell destruction.
In terms of immunology and pathomorphology, the pathogenesis of GD is thought to be related to T-helper type 2(Th2), with T-cell-mediated response and inflammatory infiltration. The major autoantigen in Graves' disease patients is thyroid stimulating hormone receptor (TSHR), and also present in a small number of patients with Hashimoto's thyroiditis(7). Specific thyroid stimulating hormone receptor autoantibody (TRAb) stimulates the formation of TSHR, leading to hyperthyroidism. Generally TRAb refers to any type of antibody that specifically interacts with TSHR. High TSHR antibody levels at diagnosis or positive TSHR antibody at cessation of treatment indicate a high probability of recurrence within 2 years. The stimulating effect of TRAb on thyroid cells may also influence the formation of TgAb and TPOAb. The level of thyroid autoantibody titer is also important, but the exact mechanism is not clear.
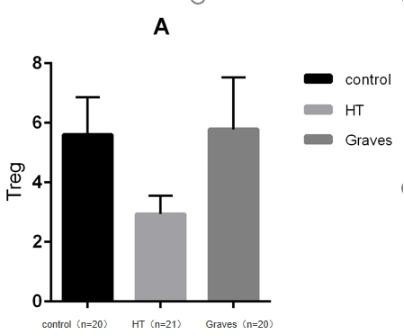
In addition to thyroid autoantibody production and abnormal thyroid hormone production, AITD is histologically involved in targeting T and B lymphocyte infiltration in the thyroid(1). Different immune cell subsets play an important role in the pathogenesis and tissue damage of AITD, among which the activation of regulatory T cells and helper T cell subsets and the imbalance of cytokines secreted by them are important links leading to the pathogenesis of AITD. The main function of regulating T cells is to inhibit the activation and proliferation of CD4 + CD8 + T cells and play a negative regulatory role in the immune response. The decreased function of regulating T cells can lead to relatively active functions of CD4 + CD8 + T cells and the imbalance of Th1/Th2, resulting in autoimmune disorders dominated by cellular immune hyperfunction(8)(9)(10). Regulatory T cells (Tregs) are primarily responsible for maintaining self-tolerance, and their protective effects have been demonstrated in animal models of several autoimmune diseases. MorrisGP et al. showed that Treg depletion enables thyroiditis induction with mouse thyroglobulin (mTg) in traditionally-resistant mice and mTg-induced, Treg-mediated tolerance protects against EAT(a model for Hashimoto’s thyroiditis) induction in genetically-susceptible mice. Meanwhile, they proposed that the phenotype of Treg is CD4 + CD25 + Foxp3 + regulatory T cells. These data demonstrate that tregs are essential in maintaining peripheral tolerance and have potential therapeutic in inducing tolerance(11). In addition, Treg cells have been linked to chronic inflammation and other autoimmune diseases such as systemic lupus erythematosus, Sjogren's syndrome and systemic sclerosis. There are many subsets of regulatory T cells classified from the origin. These subsets have different T cell receptors (TCR), and have different functions in local tissues. FOXP3 + Treg cells are generally regarded as active regulatory T cells. Although many markers have been identified that play an important role in the immunobiology of Tregs, most are not specific to Tregs. In this experiment, CD4 + CD25 + CD127lowT cells were set as Treg cells in the study.
CD25 (also known as IL-2R receptor or α-subunit of IL-2 receptor) is involved in the regulation of T cell function. It is encoded by the CD25 region on chromosome 10p15.1, highly expressed in Tregs, and mediates IL-2 signal transduction, which is essential for the survival and growth of CD25 + CD4 + Tregs(12). It is known that FOXP3 is a master regulator for Treg cell differentiation and function. Foxp3 negatively regulates the transcription of IL-2 and up-regulates the expression of IL-2R, making exogenous IL-2 a necessary condition for the survival of Treg cells. It has been shown that the signal transduction of IL-2 is particularly critical to the maintenance of Treg homeostasis and FOXP3 expression. PangdiYang et al. 's study showed that tregs can absorb IL-2 effectively due to the high expression of CD25, thus depleting IL-2 in the environment, leading to the release of granzyme and perforin, and inducing cell apoptosis. CD25 is highly expressed on the surface of treg cells and binds to IL-2 with high affinity, then JAK/STAT signaling pathway is activated, thus maintaining the expression of FOXP3 and the development, proliferation and inhibitory functions of Tregs. Similar to FOXP3 impaired mice, IL-2Rαdeficient mice showed similar lethal lymphoproliferative disease with severe autoimmunity(13). Therefore, it is hypothesized that some genetic variants in the CD25 gene predispose to autoimmune disease by impairs Treg cell function and peripheral tolerance.In fact,a case-control study from Britain reported a significant correlation between CD25 and GD(14). NTregs can inhibit multi-organ autoimmunity induced by CD4 + CD25-T cells in nude mice, and Tregs consumption in vivo increased the incidence of autoimmune diseases including thyroiditis in other resistant mouse.
Interleukin-7 (IL-7) is a crucial cytokine involved in T-cell survival and development, and CD127 expression ensures an appropriate response to IL-7 and maintains T-cell homeostasis(15)(16)(17). IT is known that FOXP3 is a key regulator of Treg cell differentiation and function, and regulatory CD4 + FOXP3 + T cells express very little CD127 on their surface,so a combination of CD4, CD25 and CD127 antibody can be used to evaluate the phenotype of Treg cells (CD4 + CD25 + CD127-T cells). CD4 + CD25 + CD127 (low) T cells express high levels of Foxp3 and exert high immunosuppression against effector T cells(18)(19). Studies have shown that the expression of CD127 is decreased on CD4 + T cells and the proportion of CD4 + CD25 + CD127 (low) T cells is increased in the peripheral blood of children with type 1 diabetes with a longer duration, and there is a good correlation between them(20). Currently, CD4, CD25 and CD127 combinations have been used to select T cells for functional research as well as for expansion and adoptive immunotherapy after expansion.
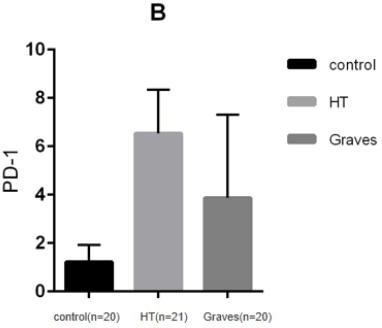
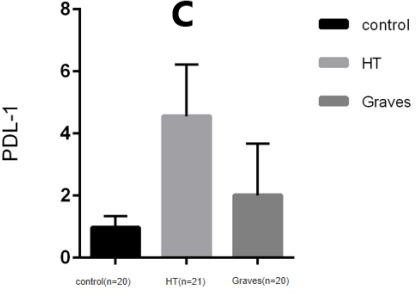
There are two independent signals in the classical T cell activation pathway, One is the specific signal provided by the interaction between the MHC-antigen (Ag) complex on antigen presenting cells (APCs) and T cell receptors (TCRs) on T cells, and the other is the non-antigen-specific signal transmitted by the co-stimulation molecules expressed on the surface of T cells and APCs cells. The costimulatory signaling pathway not only provides a critical positive second signal to initiate, enhance, and maintain the T-cell response, but also provides a critical negative second signal to limit, terminate, or impair the T-cell response. Programmed cell death-1 (PD-1)/programmed cell death ligand (PD-L1) costimulatory pathway is a negative costimulatory pathway discovered in recent years, PD-1 and its ligands PD-L1 (B7-H1, CD274) and PD-L2 (B7-DC, CD273) plays an important role in the induction and maintenance of peripheral tolerance and for the maintenance of the stability and the integrity of T cells. However, the PD-1:PD-L1/L2 pathway also mediates potent inhibitory signals to hinder the proliferation and function of T effector cells. Studies have shown that enhancing or blocking this pathway is associated with the immune process of AITD. PD-1, a type I transmembrane glycoprotein, is a co-inhibitory receptor of the immunoglobulin gene superfamily. In humans and mice,the gene encoding PD-1 is composed of 5 exons. Exon 1 encodes signal sequence, exon 2 encodes IgV-like domain, exon 3 encodes stem and TM domain, and exon 4 and 5 encodes cytoplasmic domain.It is mainly expressed in activated T cells and B cells(21). PD-1 is mainly developed in the thymus, and can also be induced to express in various hematopoietic cells by antigen receptor signals and cytokines in the periphery. As mentioned above,PD-1 has two ligands, PD-L1(B7-H1; CD274) and PD-L2(B7-DC; CD273), the difference between PD-L1 and PD-L2 lies in their different affinity for PD-1. The affinity of PD-L2 to PD-1 is three times higher than that of PD-L1, but the expression of PD-L1 is more extensive than that of PD-L2. PD-L1 can be induced to express in various hematopoietic and non-hematopoietic cells as well as various malignant cell, such as B cells, dendritic cells, macrophages, (BM-derived) mast cells and T cells, and can be upregulated further after activation. PD-L2 can be induced to express in dendritic cells, macrophages, peritoneal B cells, memory B cells and (BM-derived) mast cells. After pd-1 binds to its ligand, Intracellular tyrosine residues are phosphorylated and phosphatases are subsequently activated, then proximal TCR signal transduction is inhibited, ultimately inhibiting T cell activation, proliferation, and cytokine secretion(22)(23). Therefore, the interaction between Treg cells expressing PD-1 and PD-L1 and conventional T cells can lead to negative regulation of Treg cells and conventional T cells.
{kind=link}
{kind=link}
{kind=link}