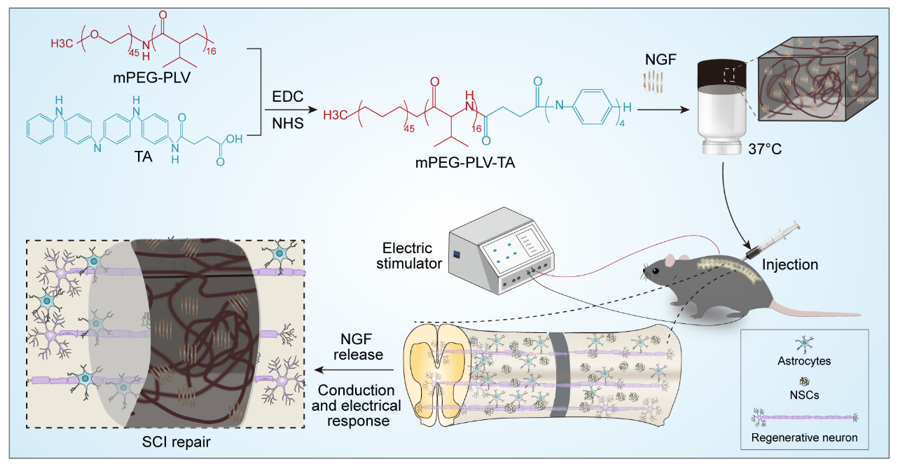
3.1. Characterization of mPEG-PLV-TA copolymer
The electroactive copolymer was synthesized by condensation crosslinking of mPEG-PLV and CTA. To confirm the structure of the electroactive copolymer, we tested 1H-nuclear magnetic resonance and Fourier transform infrared spectroscopy. All peaks in the copolymer were clearly assigned (Fig. 1A), demonstrating successful synthesis. Typical CTA absorption peaks were observed at 1603 and 1508 cm− 1 (Fig. 1B) which corresponded to the stretching vibration of quinone and the benzenoid unit in CTA chain, which indicated that CTA was successfully grafted to mPEG-PLV.
Ultraviolet–visible light (UV–vis) absorption spectra of different oxidation states of the electroactive copolymer are shown in Fig. 1C. There were three oxidation states—i.e., leucoemeraldine (LM), EM, and pernigraniline (PN). When the copolymer was gradually oxidized, the benzenoid unit was replaced by a benzene ring. In the LM state, the only one absorption peak showed at 310 nm can be related to the π–π* transition of the benzenoid unit. In the EM state, this absorption peak showed a marked blue shift and a new peak appeared at 575 nm corresponding to the excitonic transition of π–π* from benzenoid to the quinone unit [22]. When the copolymer in the EM state was doped with HCl (1 M), new characteristic absorption peaks appeared at 430 nm and > 800 nm, reflecting the formation of the EMS state and demonstrating the conductivity of the copolymer after doping [23]. Figure 1D showed the UV–vis spectrum of the gradual oxidation of the mPEG-PLV-TA electroactive copolymer. When ammonium persulfate was gradually added to the DMF solution of the electroactive copolymer, the intensity of the absorption peak at 575 nm gradually increased; after reaching a maximum value, the intensity gradually diminished and showed a blue shift as the degree of oxidation increased.
This redox process was also be observed in the current–voltage curve of the copolymer (Fig. 1E). The first pair of redox peaks reflected the transition of TA between EM and LM states, whereas the second pair of peaks corresponded to the transition of TA from LM to PN states.
3.2. Gelation ability and internal structure
Different concentrations of copolymer were dissolved in PBS at 4°C to obtain homogeneous solutions. The temperature was gradually increased and changes in the state of the solution were examined. As the copolymer concentration increased from 4.0 to 8.0 wt%, a stable gel was formed while the temperature of the sol-gel transition decreased from 33.3°C to 5.0°C (Fig. 1F). Considering the issues of injectability and gelation time during implantation, we selected the 6.0 wt% copolymer solution to prepare electroactive hydrogels for animal experiments. To determine whether the internal structure of the hydrogel was suitable for SC regeneration, we examined the ultrastructure by SEM. The hydrogel had uniform pores with a diameter of 30–70 µm (Fig. 2A), which can serve as channels for tissue fluid, exchange of metabolic substances, drug release, and axon growth.
3.3. NGF release and hydrogel degradation
Long-term nutrient deficiency after SCI impedes the speed and ultimate extent of tissue repair; this problem can potentially be overcome using a scaffold loaded with nutrients, and we therefore investigated whether the hydrogel can efficiently release NGF, which was substituted with lysozyme in this experiment. We loaded 20 mg/mL lysozyme into the electroactive hydrogel and observed the release kinetics [21]. Lysozyme encapsulated in the hydrogel showed burst release (48.28%±2.01%) in the first 4 days (Fig. 2B), which may have been due to its leaching out of the hydrogel with water. To simulate the release of lysozyme in the body with hydrogel degradation, elastase (2 mg/L) was added to PBS and release kinetics were monitored [24]. After adding elastase, about 87.02%±3.05% of the lysozyme was released from the hydrogel within 24 days, suggesting that the hydrogel can release loaded NGF in vivo over a sufficiently long period of time to allow nerve cell growth after SCI.
We evaluated the in vitro degradation of the hydrogel using samples in PBS alone or PBS containing elastase. Degradation was accelerated in the presence of elastase, with a hydrogel mass loss of > 80.3%±1.86% within 30 days (Fig. 2C). These results demonstrate that the hydrogel is degraded at a moderate rate, which can benefit long-term tissue repair.
To further assess the degradation process and biocompatibility, the hydrogel was implanted under the skin of Sprague–Dawley rats. The hydrogel slowly degraded over a period of 4 weeks at a slightly faster rate than in vitro (Fig. 2D). We performed H&E staining of the skin contacting the hydrogel obtained at different time points and found that acute inflammation occurred after the hydrogel was injected and was most prominent on day 7. Over time, the inflammatory response gradually declined and there was no inflammation on day 28.
3.4. Biocompatibility and cytotoxicity assessment in vitro
To evaluate the in vitro cytotoxicity of the electroactive hydrogel, we carried out live–dead cell and MTT assays. PC12 cells were cocultured with hydrogel in mixed calcein-AM and PI dye solution; after 5 days, the cells were proliferating with no evidence of cell death (Fig. 3A). In the MTT assay, when the concentration of the hydrogel suspension increased from 0 to 500 µg/mL, cell viability was in the range of 97.38%±6.11–100%±3.12% after 24 h (Fig. 3C) and 90.40%±1.22–100%±3.48% after 48 h (Fig. 3D). Thus, the electroactive hydrogel has excellent biocompatibility and can be safely used for tissue repair.
3.5. Axon growth and NSC differentiation in vitro
CPs can stimulate axon growth to promote nerve repair [25]. To investigate whether it can perform this function, the electroactive hydrogel was cocultured with PC12 cells and the length of axons emerging from the cells was analyzed [26]. In the Gel + NGF and Gel + ES groups, only a few cells showed axon-like protrusions; in contrast, most cells in the Gel + NGF + ES showed neurite outgrowth (Fig. 3B). The average neurite length in the Gel + NGF + ES group was 136.30 µm, which was significantly longer than that in the control (25.48 µm), Gel + NGF (38.64 µm), and Gel + ES (45.12 µm) groups (Fig. 3E). These results demonstrate that while the hydrogel by itself has limited ability to stimulate axon growth in nerve cells, the effect can be enhanced by ES and NGF.
In addition to promoting axon growth, replacing dead neurons in the SC is an important strategy for restoring motor function. To verify whether the hydrogel can promote a large-scale differentiation of NSCs into neurons, NSCs were cocultured with the hydrogel and immunolabeling was performed to assess their differentiation status after 1 and 7 days. We found that the hydrogel induced the differentiation of NSCs into neurons and that the effect was potentiated by ES. This was accompanied by changes in cell morphology: neurites of a single neuron extended in all directions, forming an extensive network with other neurons (Fig. 4A). During this process, there were no changes in the expression level of nestin, a marker of NSCs, in any treatment groups (Figs. 4B and 5B). However, there was a significant increase in Tuj1 expression in all groups from day 1 to 7; the average fluorescence intensity on day 7 was 8.73 in the Gel + NGF + ES group, which was significantly higher than that in the control (5.21), Gel + NGF (6.37), and Gel + ES (7.02) groups (Fig. 4C). In contrast, the hydrogel had an inhibitory effect on NSC differentiation into astrocytes (Fig. 5A): the average fluorescence intensity of GFAP was lower in the Gel + NGF + ES group (4.01) on day 7 than in the control (7.58), Gel + NGF (6.95), and Gel + ES (5.24) groups (Fig. 5C). These results suggest that the hydrogel inhibits astrocyte proliferation and may prevent the formation of glial scars.
3.6. Recovery of motor function and nerve conduction
The restoration of motor function is the most important index for gauging SC repair. For the purpose of investigate the ability of the hydrogel to promote functional recovery, the footprint test was conducted and BBB score was evaluated in rats with SCI. At postoperative week 12, the sham group showed a strong and stable normal gait in the footprint analysis, whereas injured rats showed a decreased stride length and increased sway distance with severe toe dragging (Fig. 6A). The treatment groups showed varying degrees of gait recovery: rats in the Gel + NGF + ES group had a stride length (10.55 ± 0.21 cm) that was closer to that of the sham group (11.38 ± 0.22 cm) than the Gel + NGF (9.13 ± 0.29 cm) and Gel + ES (9.48 ± 0.21 cm) groups, indicating that the recovery of motor function was greatest with the hydrogel loaded with NGF combined with ES (Fig. 6C). The same trend was observed for sway distance (2.63 ± 0.13 cm in the Gel + NGF + ES group vs 2.28 ± 0.13 cm in the sham group) (Fig. 6D). Abnormal gait may be caused by decreased muscle tone, cerebellar dysfunction, impaired peripheral nerve function, or musculoskeletal abnormalities [27]. In this study, the degree of gait recovery was different, which could be due to the different degrees of regeneration and conduction of nerve cells in the gray and white matter, respectively; alternatively, inconsistent muscle tension caused by atrophic lower limb muscles may account for the observed differences.
To monitor the recovery of motor function of rats with SCI, we determined the BBB score in multiple different time points. The right hind limb of SCI model rats was paralyzed immediately after the operation, and the BBB score was close to 0 (Fig. 6B); even after 12 weeks, the score did not exceed 4 points, indicating that there was very little recovery after injury. In the treatment groups, the BBB score gradually increased over time; starting from week 4 after surgery, the fraction of Gel + NGF, Gel + ES, and Gel + NGF + ES groups are significantly higher scores than the SCI model group and after 12 weeks, the BBB score of the Gel + NGF + ES group (13.8 ± 0.84) was higher than those of the other treatment groups.
We also evaluated the recovery of nerve conduction by measuring MEP. The amplitude of the MEP was reduced by SCI while the latency was increased, and there was no improvement in these parameters over time (Fig. 6E, F). After 12 weeks, the amplitude was increased in all treatment groups and the latency was shortened. The amplitude of the MEP was obviously higher in the Gel + NGF + ES group (2.47 ± 0.14 mV) than in the Gel + NGF (1.62 ± 0.08 mV) and Gel + ES (1.63 ± 0.11 mV) groups. Similarly, the latency was shorter in the Gel + NGF + ES group (3.73 ± 0.18 ms) than in the Gel + NGF (6.81 ± 0.13 ms) and Gel + ES (4.3 ± 0.1 ms) groups. Taken together, these results demonstrate that the NGF-loaded electroactive hydrogel combined with ES promoted the recovery of motor function in rats after SCI.
3.7. Tissue repair in the SC
We carried out histologic and ultrastructural analyses to confirm the repair of SC tissue. Rats were sacrificed 12 weeks post surgery and the SC was completely dissected. H&E staining of tissue sections revealed that there was no new tissue growth in the SCI model group (Fig. 7A). However, the Gel + NGF + ES group had smallest cavity area corresponding to the site of injury and more new tissue than the other treatment groups (Fig. 7B). A closer examination of SC ultrastructure by TEM indicated that compared to the sham group, SCI model rats had scar tissue hyperplasia and demyelination, with no obvious myelin sheath or axon regeneration, whereas myelinated axons were observed in the Gel + NGF, Gel + ES, and Gel + NGF + ES groups (Fig. 7C, D). These results demonstrate that the hydrogel with NGF and ES can promote nerve cell growth and SC tissue recovery.
3.8. Differentiation of NSCs into neurons and inhibition of glial scar formation in vivo
To determine whether the electroactive hydrogel loaded NGF combined with ES can facilitate the differentiation of NSCs into neurons in vivo, we performed double immunofluorescence labeling of SC tissue specimens with antibodies against nestin and Tuj1. There were no newborn neurons (Tuj1-positive cells) in the SCI model group, but they were observed in rats treated with Gel + NGF, Gel + ES, and Gel + NGF + ES (Fig. 8A). Notably, nestin and Tuj1 showed different degrees of expression and colocalization in areas of new tissue, suggesting that NSCs migrated from intact parts of the SC to the site of injury before differentiating into neurons. A semiquantitative analysis illustrated that the rate of neurogenesis in the Gel + NGF + ES group was 1.2 and 3.1 times higher than that in Gel + NGF and Gel + ES groups, respectively (Fig. 8B, C).
Astrocytes are the dominant type of glial cell in the central nervous system and their continuous activation and hyperproliferation after SCI can lead to the formation of glial scars, which can prevent SC regeneration. Double immunolabeling of GFAP and Tuj1 in SC tissue sections proved that GFAP expression was higher in SCI model rats than in sham group (Fig. 9A, C) and higher in the Gel + NGF group than in the Gel + ES and Gel + NGF + ES groups. This may be related to the fact that ES inhibits glial scar formation. In addition to being expressed by immature neurons, Tuj1 is also an axon marker. Tuj1 expression was observed in all treatment groups, and the level in the Gel + NGF + ES group was significantly higher than that in the Gel + NGF and Gel + ES groups (Fig. 9A, B). Thus, the electroactive hydrogel loaded with NGF can promote axon regeneration in the injured SC, with ES enhancing this effect.
{kind=link}