Symptoms of Leaf Sheath after Inoculation
The symptoms of leaf sheath after inoculation are shown in Figs.1. After inoculation for 24 h, evident brown spots appeared on the leaf sheath of Yueguang; 48 h after inoculation, they were grey. These spots appeared on the leaf sheath of Shennong 9819 at 36 h after inoculation. The expanded area of the spot of Yueguang was significantly larger than that of Shennong 9819 at 72 h after inoculation.
The inoculated leaf sheath was decolorized by chloral hydrate, stained with aniline blue, and observed under a light microscope. At 12 h after inoculation, hyphae were observed in the leaf sheaths of both cultivars, and more hyphae were found in the leaf sheath of Yueguang than in Shennong 9819. At 24 h after inoculation, infection cushions appeared in the leaf sheaths of both cultivars. The number and density of the infection cushions of Yueguang were greater than those of Shennong 9819 (Figs.2).
RNA-seq Results of Transcriptome Samples
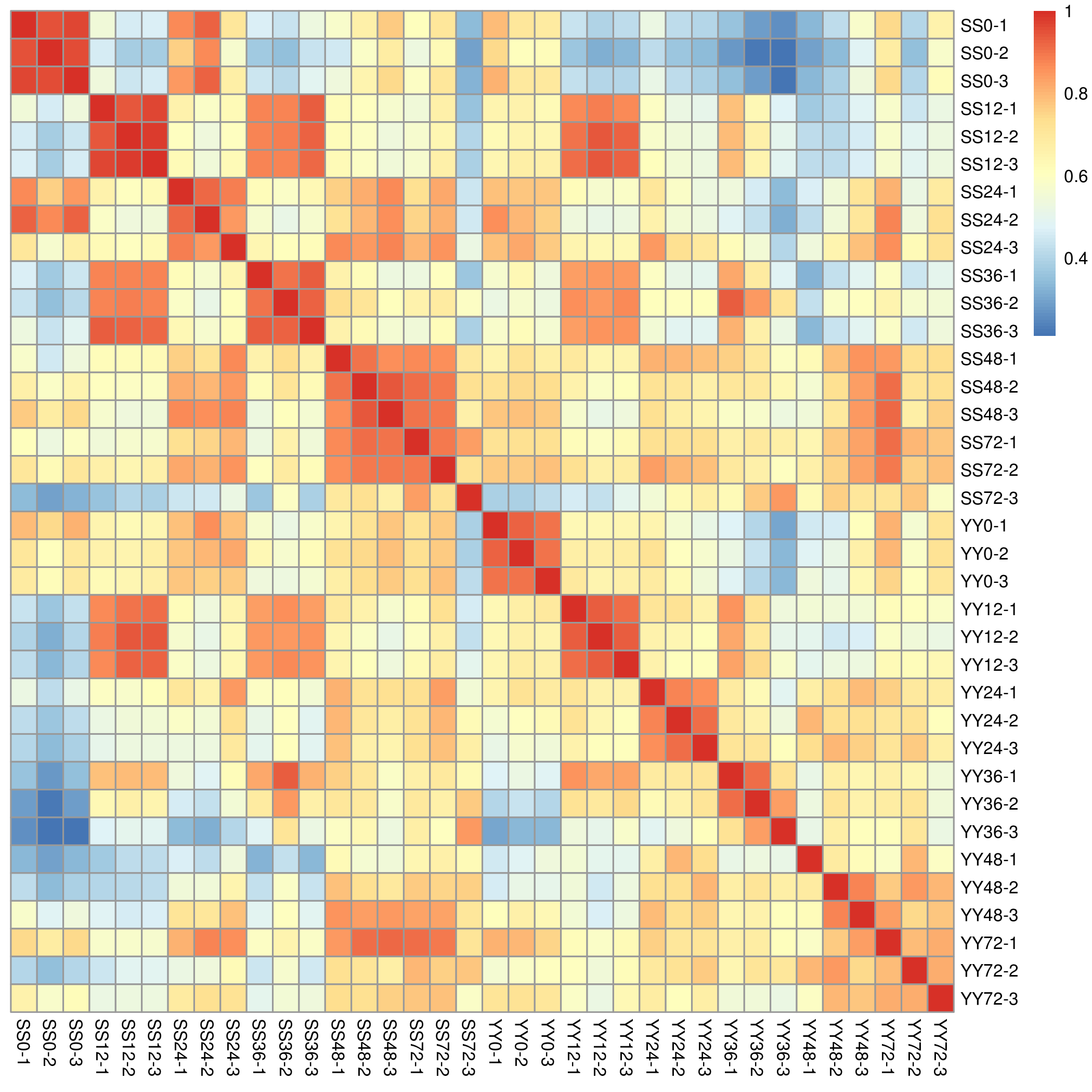
To study the changes in gene expression of the leaf sheath of Shennong 9819 and Yueguang at the initial infection stage of R. solani, we used high-throughput sequencing technology to measure the transcription in rice leaf sheaths after inoculation. The transcriptome analysis of each cultivar included five time points with three biological repeats at each time point. A total of 342.24 Gb clean data was obtained from 36 samples; the clean data of each sample reached 8.08 Gb, and the percentage of Q30 base was 92.49% or more (Table S1). A total of 2,294,868,004 single-end clean reads (total records) were obtained after pre-processing the reads (Table S2). The clean reads of each sample were sequenced with the designated reference genome, and the efficiency of alignment ranged from 80.43% to 92.33%. The correlation analysis among the samples showed that the three repeats of the two cultivars had a high correlation (Figs.3).
Differential Gene Analysis of Leaf Sheath after inoculation
To determine which gene expression had changed and the stage of these changes, we counted the number of different genes between the two rice cultivars at each time point after inoculation with R. solani (Table 1). A total of 2275 differentially expressed genes (DEGs) were identified in our study (Table S3). After inoculation with R. solani, the number of upregulated genes in the leaf sheath of Yueguang was higher than that of Shennong 9819 at all inoculation time points.
Table 1
Statistics of differentially expressed genes
|
DEG Set
|
DEG Number
|
up-regulated
|
down-regulated
|
|
SS12
|
2,403
|
1,242
|
1,161
|
|
SS24
|
190
|
129
|
61
|
|
SS36
|
2,127
|
815
|
1,312
|
|
SS48
|
790
|
658
|
132
|
|
SS72
|
1,120
|
818
|
302
|
|
YY12
|
2,817
|
1,419
|
1,398
|
|
YY24
|
3,392
|
1,983
|
1,409
|
|
YY36
|
4,873
|
2,438
|
2,435
|
|
YY48
|
1,303
|
661
|
642
|
|
YY72
|
1,766
|
881
|
885
|
At 12 h, 2,403 DEGs (1,242 upregulated and 1,161 downregulated) were identified in the leaf sheath of Shennong 9819, whereas 2,817 DEGs (1,419 upregulated and 1,398 downregulated) were identified in Yueguang, indicating that the Yueguang sheath was more susceptible to R. solani than Shennong 9819; the infection pressure on Yueguang plants was thus higher than that on Shennong 9819. From 12 to 72 h, the number of DEGs in the Yueguang leaf sheath was higher than Shennong 9819. The number of DEGs in the Yueguang leaf sheath was highest at 36 h (4,873 DEGs; 2,438 upregulated, 2,435 downregulated) after inoculation. The data show that the number of DEGs in the susceptible cultivar was higher than that in the resistant cultivar.
The DEGs at 12, 24, 36, 48, and 72 h after inoculation of R. solani were analysed for the two cultivars. The DEGs related to the infection response of R. solani in rice were further analysed.
In this study, the DEGs of the two cultivars at the same time point (SS12-YY12, SS24-YY24, SS36-YY36, SS48-YY48, and SS72-YY72) were compared (Fig.1), and the number of DEGs for each cultivar at different time points (SS12-SS24-SS36-SS48-SS72 or YY12-YY24-YY36-YY48-YY72h) were also compared (Fig.2). As shown in Fig.1, 1,347 and 1,269 DEGs were identified in the two cultivars at 12 h and 36 h after inoculation, respectively, and the number of these DEGs was higher than that at other time points.
In the leaf sheaths of Shennong 9819, 12 DEGs were continuously expressed at 12, 24, 36, 48, and 72 h after inoculation, including four upregulated genes and seven downregulated genes. At 12 h, 832 DEGs were identified (575 upregulated and 268 downregulated). There were 23 DEGs (15 upregulated and 14 downregulated) discovered at 24 h, and 693 DEGs (284 upregulated and 419 downregulated) were identified at 36 h. In the leaf sheath of Yueguang, 269 DEGs were expressed at 12, 24, 36, 48, and 72 h after inoculation (132 upregulated and 137 downregulated). At 12 h, 537 DEGs (417 upregulated and 172 downregulated) were identified, and 925 DEGs (619 upregulated and 359 downregulated) were identified at 36 h. At 36 h, 1,251 DEGs were identified (612 upregulated and 645 downregulated). In conclusion, the number of DEGs which were continuously expressed in the sheath of Yueguang was higher than that in Shennong 9819 after inoculation.
Gene Ontology (GO) Analysis of Differentially Expressed Genes
GO annotation was used to classify the enriched DEGs between the control and inoculation treatments. The results showed that these enriched DEGs were involved in many biological activities (Fig.3).
After annotating the GO database, all the DEGs were classified into three main categories: biological process, molecular function, and cellular component. In the biological process, “metabolic process”, “cellular process”, “single-organism process”, “response to stimulus”, and “biological regulation” were the five processes with the highest degree of DEGs enrichment. In terms of molecular function, most DEGs were concentrated in two processes: “catalytic activity” and “binding”. For the cellular component, the five most common processes were “cell”, “membrane”, “organelle”, “organelle part” and “cell part”.
The results showed that the GO terms in different cultivars showed completely different patterns of expression. Taking “response to stimulation” as an example, in the leaf sheath of Shennong 9819, the number of DEGs related to this process reached the maximum at 12 h. The number of DEGs was the least at 24 h after inoculation, indicating that plants responded most strongly to external stimuli at 12 h and were closest to the uninoculated state at 24 h. The results showed that the plants could repair themselves in response to external stimuli; at 36 h, the number of DEGs increased again, and at 48 h, the number of DEGs decreased again. Shennong 9819 exhibited a fluctuating response to external stress. However, the number of DEGs related to the process of “response to stimulation” increased rapidly in the leaf sheath of Yueguang after inoculation, reached a maximum at 36 h and then decreased sharply. The genes related to this process in the leaf sheath of Yueguang were always active after inoculation. The results showed that Shennong 9819 and Yueguang had different resistance patterns to R. solani infection, and the genes in the leaf sheath of Yueguang were always in a higher activity state.
Analysis of Metabolic Pathways of Two Rice Cultivars after Inoculation
To further study the specificity of pathways affected by R. solani infection in Shennong 9819 and Yueguang, Kyoto Encyclopaedia of Genes and Genomes (KEGG) enrichment analysis was performed on the upregulated genes (|log2fc>1|, FDR < 0.05) at different times after inoculation (Table S4 and S5). The results showed that alanine, aspartate and glutamate metabolism was significantly enriched in both cultivars at 12 h after inoculation, although Shennong 9819 was more significant. The phenylalanine metabolism, plant hormone signal transduction, tropane, piperidine and pyridine alkaloid biosynthesis pathways were significantly enriched in Shennong 9819 at 12 and 36 h after inoculation; however, no significant difference was found in Yueguang. The tyrosine metabolism and isoquinoline alkaloid biosynthesis pathways were significantly enriched in Shennong 9819 at 24 and 36 h after inoculation, but not in Yueguang. At 36 h after inoculation of Shennong 9819 and Yueguang with R. solani, glycine, serine and threonine metabolism and beta-alanine metabolism pathways were significantly enriched, whereas Yueguang was not.
Similarly, some pathways were specifically enriched in Yueguang. For example, ascorbate and aldarate metabolism pathway was significantly enriched at 12 and 24 h after inoculation in Yueguang. The linoleic acid metabolism pathway was significantly enriched 24 h after inoculation in Yueguang. Valine, leucine and isoleucine degradation pathway was significantly enriched at 24 and 36 h in Yueguang after inoculation. The arginine biosynthesis and 2-Oxocarboxylic acid metabolism pathways were significantly enriched at 36 h in Yueguang after inoculation. The propanoate metabolism pathway was significantly enriched at 24, 36, 48, and 72 h in Yueguang after inoculation. The above pathways were not significantly enriched at any time in Shennong 9819 after inoculation. At different times after inoculation, the upregulated genes of the two cultivars had specific enrichment pathways, suggesting that the resistance mechanisms of the two cultivars might be different.
To identify potential regulatory genes closely related to the phenylalanine metabolism pathway, DEGs involved in phenylalanine metabolism were identified by comparing the two cultivars. At 12 and 36 h, the expression levels of genes related to phenylalanine metabolism in the Shennong 9819 leaf sheath were significantly higher than those in Yueguang. The difference between the two cultivars is shown in the heatmap (Fig.4). Through gene analysis, a series of PAL genes was activated after inoculation with R. solani: OsPAL1 (LOC_Os02g41630), OsPAL2 (LOC_Os02g41650), OsPAL3 (LOC_Os02g41670), OsPAL4 (LOC_Os02g41680), OsPAL6 (LOC_Os04g43800), and OsPAL9 (LOC_Os12g33610). In this study, the expression of OsPAL1 and OsPAL6 in the leaf sheath of Shennong 9819 was higher than that in Yueguang (Fig.5).
Furthermore, we compared the DEGs involved in plant hormone signalling pathway between the two cultivars. We found that the expression of genes closely related to plant hormone signal transduction in the leaf sheaths of the two cultivars was similar to that of phenylalanine metabolism. At 12 and 36 h, the expression levels of genes related to plant hormone signal transduction in Shennong 9819 leaf sheaths were significantly higher than those in Yueguang (Fig.6). The resistance-related protein kinases OsSAPK9 (LOC_Os12g39630) [36] and OsSAPK10 (LOC_Os03g41460) [37] were upregulated in the leaf sheaths of Shennong 9819, and their expression levels were higher than those of Yueguang (Fig.5). In addition, OsNPR1 (LOC_Os01g09800), a positive regulator related to resistance, was also detected and upregulated in both cultivars (Fig.5). In the early stage of infection with R. solani, the two cultivars initiated similar pathways in response to stimulation; however, the expression of resistance-related genes may be different in time and expression levels.
As previously mentioned, the two cultivars participated in similar metabolic pathways after inoculation with R. solani; however, the upregulated differential expression pathways were different. Among the pathways related to disease resistance, plant hormone signal transduction and phenylalanine metabolism pathways were significantly enriched in Shennong 9819, except in Yueguang. However, ascorbate and aldarate metabolism, and linoleic acid metabolism were significantly enriched in Yueguang, though not in Shennong 9819. It is necessary to further study the differentially expressed genes in the metabolic pathway to understand the different resistance mechanisms of the two cultivars after inoculation with R. solani.
Validation of DEGs by Quantitative RT-PCR (qRT-PCR)
Real-time quantitative PCR (qRT-PCR) was used to validate RNA-seq data. OsPR1b and other genes were selected for further validation [38]. The Ct values obtained by qRT-PCR were normalised. The fold change in gene expression of the two rice cultivars inoculated with R. solani was calculated. The results showed that the expression trend of qRT-PCR was consistent with that of the RNA sequence, indicating that Illumina data were relatively reliable (Figs.4).
{kind=link}