Biological diversity comprises community composition, structure, and function [38]. Interactions between soil microorganisms and other organisms influence nutrient cycling, which plays an essential role in soil condition, quality, and health [13, 39, 40]. The rhizosphere is a functional interface for materials exchange between plants and soil ecosystems. Plants assimilate CO2 during photosynthesis and transport some photosynthetic products to their underground parts, promoting the growth and metabolism of soil microorganisms, which in turn transform organic nutrients in the soil into inorganic forms for absorption and utilization by plants [41]. With the recent emergence of transgenic plants, the impact of their cultivation on the structure and function of the rhizosphere microbial community has become a concern [16, 42, 43]. The structural diversity of the soil microbial community is an important index for evaluating the effects of GM on the soil ecological environment.
Changes in soil MBC, MBN, and MBP content in WT and Cry1Ah1-modified poplars
Soil microbial biomass is an essential parameter for the assessment of active soil nutrients and a sensitive indicator of environmental change in terrestrial ecosystems [44–46]. MBC, MBN, and MBP participate in the ecosystem cycling of carbon, nitrogen, and phosphorus [47, 48]. To date, the effects of GM plants on MBC, MBN, and MBP have not been reported. Therefore, in the present study, we examined the impact of field-cultivated GM plants on MBC, MBN, and MBP content to study the relationship between GM plant growth and the transformation of carbon, nitrogen, and phosphorus in natural soil.
As a part of active soil carbon, MBC is the driving force of soil organic matter decomposition, which is closely related to the cycling of soil elements. We found that MBC content was significantly higher in Cry1Ah1-modified varieties than in WT poplars. MBC content decreased in Cry1Ah1-modified poplars grown in the field, which may have affected the growth, metabolism, and structure of soil microorganisms. As an essential source of active soil nitrogen, MBN plays a vital role in regulating the supply of soil nitrogen [49]. MBP is the most active component of soil organic phosphorus, which governs the mineralization and fixation of soil phosphorus. MBP is an important source of available soil phosphorus, which reflects the capacity and turnover intensity of active soil phosphorus [50, 51]. Our results showed that Cry1Ah1-modified poplars altered soil MBN and MBP content, at least during the study period, affecting the capacity of soil microorganisms to metabolize carbon, nitrogen, and phosphorus. Plants can alter the available or bioaccessible concentrations of soil elements directly through absorption and removal or indirectly through plant residue decomposition. Total concentrations of soil elements are generally not affected by plants [52, 53]. However, we found that available soil phosphorus, MBN, and MBP content were decreased, whereas MBC content was significantly increased in Cry1Ah1-modified poplars. This result can be used to guide future poplar breeding. Although Cry1Ah1 transformation can improve poplar insect resistance, transgenic poplars grown in fields may be affected by MBC, MBN, and MBP, which will eventually affect the growth of plants.
The soil MBC/MBN ratio can reflect soil microbial community structure. Generally, the MBC/MBN ratio is about 5:1 for bacteria, 6:1 for actinomycetes, and 10:1 for fungi [54–58]. Based on our results, the MBC/MBN ratio for WT poplars in our study site was about 4.6 ± 0.3, indicating that bacteria may play a dominant role in determining MBC and MBN contents. However, the MBC/MBN ratio for Cry1Ah1-modified poplars was about 9.2 ± 1.7, suggesting that microbial fungi in Cry1Ah1-modified poplars participate widely in soil microenvironment regulation.Xu et al. [32] performed a systematic analysis of MBC, MBN, and MBP in the global terrestrial ecosystem. They reported mean values of MBC/MBN, MBC/MBP, and MBN/MBP ratios of 7.6, 42.4, and 5.6, respectively. In the present study, the MBC/MBP and MBN/MBP ratios for WT poplars were 63.2 ± 3.1 and 13.6 ± 0.9, respectively, higher than those reported for the global terrestrial ecosystem. This discrepancy may be due to the low nitrogen and phosphorus content in the northern Jiangsu plain, resulting in lower MBN and MBP contents, and consequently lower MBC/MBP and MBN/MBP ratios. Compared with WT poplars, Cry1Ah1-modified poplars showed higher soil MBC/MBN, MBC/MBP, and MBN/MBP ratios; thus, Cry1Ah1 transformation may directly affect the growth of soil microorganisms or inhibit soil microbial activity, thereby affecting the metabolism of MBC, MBN, and MBP.
Soil microbial biomass is an essential biological fertility index used to evaluate soil quality [59, 60]. Nitrogen and phosphorus deficiency generally have adverse effects on soil microbial reproduction and metabolic activity, as well as on crop growth and nutrient uptake [27, 61, 62]. The lower MBN and MBP contents in Cry1Ah1-modified poplars suggested that soil microorganisms were under nutrient deficiency stress due to low metabolic efficiency and more significant release of heat and CO2 during metabolism, significantly reducing soil quality. Long-term nitrogen and phosphorus deficiency can exhaust stores of soil phosphorus, forcing the release of nitrogen and phosphorus from soil microorganisms and forming soil available nitrogen and phosphorus for absorption and use by plants. Fertilization is one approach for overcoming these adverse effects. Long-term application of inorganic fertilizers containing nitrogen and phosphorus will result in the assimilation of more nitrogen and phosphorus into microorganisms, thereby improving soil MBN and MBP. Long-term application of organic fertilizer can also speed up the decomposition of organic carbon sources by microorganisms, and more substantial assimilation rates can increase soil nutrient contents and ensure higher microbial biomass [63–66].
Effects of Cry1Ah1 expression on native rhizosphere communities
Bt protein confers strong herbicide resistance as a dominant trait of GM crops; it has been widely used in transgenic breeding to achieve herbicide-resistant plants. Xu et al. [2] showed that field-planted Cry1Ah1-modified poplars had strong herbicide resistance. With increasing numbers of GM crops worldwide, the environmental and ecological impact of GM crop cultivation has raised concerns globally. Some studies have shown that GM crops have serious negative effects on biodiversity and may pose a threat to the environment [67]. Whether GM crops affect soil microbial composition, structure, and function have become a widely studied question in the fields of ecology and food safety [68]. The plant rhizosphere is a dynamic microenvironment in which many factors, such as plant species [69], soil type [70], and root location [71], affect the composition and structure of microbial communities around plant roots [72]. Therefore, to avoid the interference of these factors on the examination of soil microbial communities in the present study, we selected poplar trees planted in a single location and collected samples simultaneously. Such a design can effectively avoid the influence of other factors on the results, focusing only on the effects of poplar type (NT or Cry1Ah1-modified) on the soil microbial community.
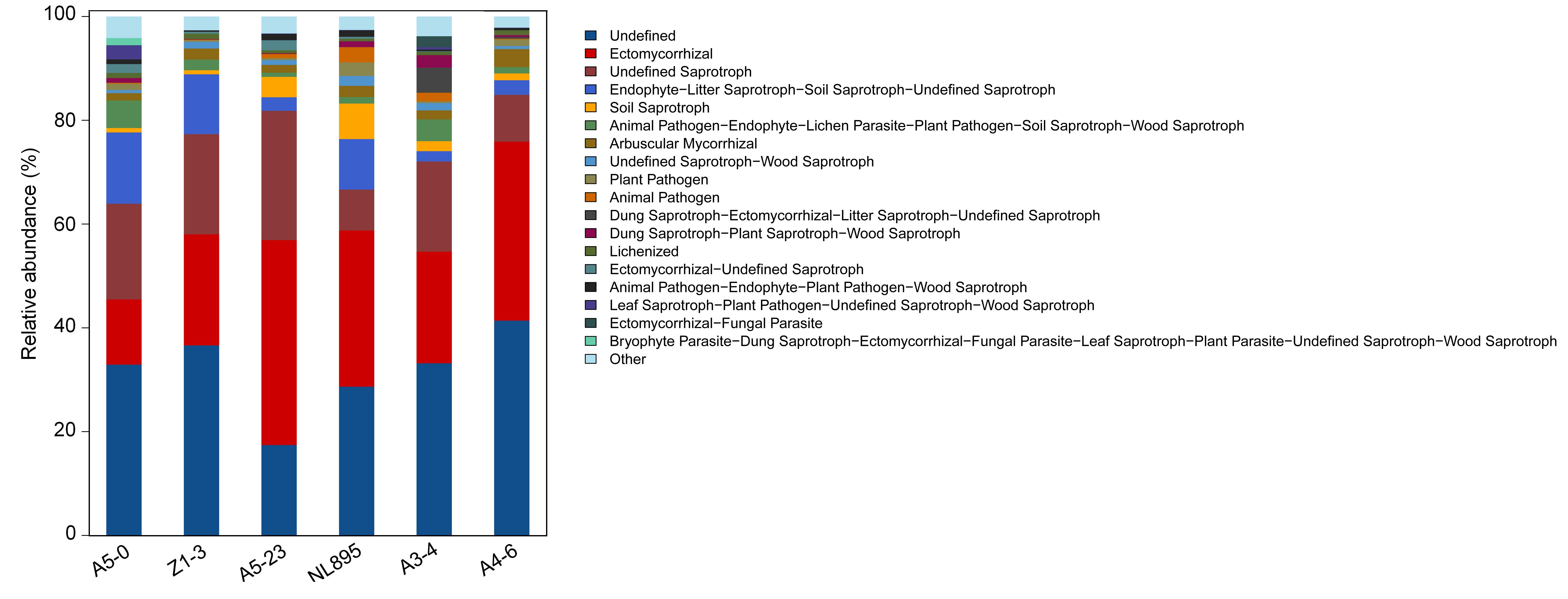
Many studies have explored the relationship between soil biodiversity and the ecological safety of transgenic plants. The high insect resistance of Bt-maize makes it an important transgenic crop, and it has been found not to affect soil microbial communities [73] or rhizosphere communities [6]. Field-cultivated Bt transgenic cotton has also been found to have no significant effect on rhizosphere communities compared with WT cotton [74]. Similarly, Cry1Ac-sugarcane was found to have no impact on rhizosphere microbial diversity or enzyme activity compared with NT sugarcane within a single crop season [75]. In addition, no persistent or adverse effects on the rhizosphere bacterial community population were detected between WT and Bt-modified rice [76]. However, Li et al. [15] reported that Bt transgenic rice had the potential to change bacterial community composition, but not fungal abundance or community structure. Thus, we conclude that most Bt-modified crops have negligible effects on soil microbial communities. In the present study, cluster heatmaps were generated from rare OTUs to show the microbial diversity and abundance of each group based on combined samples and dominant microbial genera. The results of this analysis revealed no significant difference in the structure or population of the root-associated microbial community between transgenic and NT poplars. However, the relative abundance of highly abundant bacteria, including Proteobacteria, Acidobacteria, and Actinobacteria, was affected in Cry1Ah1-modified poplars. Both the taxonomic diversity and structure of rhizosphere fungal communities were similar among high- and low-abundance fungi between Cry1Ah1-modified and NT poplars. Based on these findings, we conclude that rhizosphere microbial populations were not affected by Cry1Ah1-modified poplar cultivations, although some highly abundant bacteria in root-associated soil differed between NT and Cry1Ah1-modified poplars.
Effects of Cry1Ah1 expression on soil nitrogen and phosphorus in the microbial community
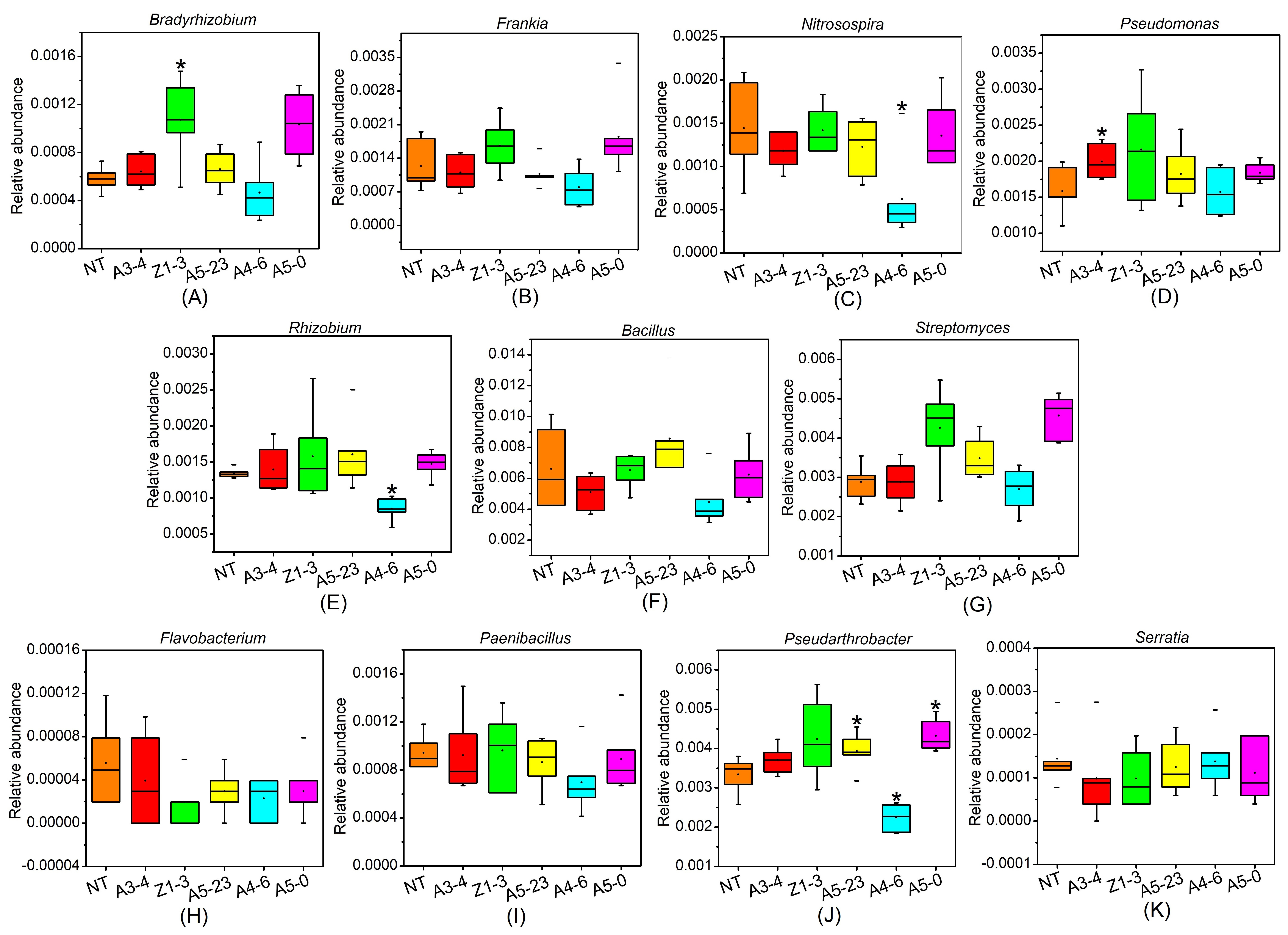
Metagenome sequencing data allowed us to identify close relationships between bacteria and nitrogen cycling based on changes in MBN content. At the genus level, four groups of nine genera of bacteria related to nitrogen cycling were isolated from rhizosphere samples. Among these, Bradyrhizobium, Rhizobium, and Frankiafour were identified as nitrogen-fixing bacteria; Nitrosospira as involved in nitrification; Pseudomonas, Ralstonia, Burkholderiales, Bacillus, and Streptomyces as denitrifying bacteria; and Bacillus and Pseudomonas as contributing to ammoniation. The relative abundances of bacteria related to nitrogen cycling decreased in the order denitrifying bacteria > nitrogen-fixing bacteria > ammonifying bacteria > nitrifying bacteria. Among these, Bacillus was the dominant genus, accounting for more than 30% of the total nitrogen-metabolizing bacteria, whereas Burkholderiales and Ralstonia accounted for smaller proportions. Our metagenome sequencing results showed that Cry1Ah1 expression resulted in no significant changes in nitrogen cycling bacterial community structure and composition, except for the relative abundance of Bradyrhizobium in line Z1-3, Nitrosospira in line A4-6, and Pseudomonas in line A3-4 (Supplementary Fig. 6). Bacillus can transform substances into nutrients needed by plants by dissolving phosphorus, fixing nitrogen, and resolving potassium, promoting plant growth. Bacillus can also affect the reproduction of some available flora, strengthening soil fertility and improving plant growth. In this study, Bacillus was the dominant genus of bacteria in NT and Cry1Ah1-modified poplars. Although the difference was not significant between NT and Cry1Ah1-modified poplars, Bacillus abundance in Cry1Ah1-modified poplars showed a downward trend, perhaps due to minor changes in bacteria related to nitrogen cycling, which can affect soil nitrogen metabolism and eventually lead to a decrease in rhizosphere MBN content.
Phosphorus is a nutrient element that forms an important component of nucleotides, phospholipids, and ATP; it participates in the physiological and biochemical processes of energy metabolism, carbohydrate metabolism, material transformation, and transportation in plants, thereby playing an essential role in growth and development [77, 78]. The phosphorus cycle is a typical depositional cycle in which phosphorus accumulates among soil, plants, and microorganisms [79]. Phosphorus exists in the soil in both insoluble or insoluble forms, and plants can utilize only 20% of them. Due to its low availability and utilization rate, phosphorus is a key element limiting plant growth [80]. Inorganic phosphorus forms can be absorbed and utilized by plants following transformation from organic phosphorus by microorganisms. This process depends on the population structure and quantity of phosphate-solubilizing microorganisms [81, 82]. In the present study, four groups of eight genera of bacteria connected with phosphorus cycling were isolated from the rhizosphere: Bacillus, Pseudomonas, Pseudarthrobacter, Flavobacterium, Paenibacillus, Bradyrhizobium, Serratia, and Streptomyces. All were found to be involved in organophosphorus metabolism. Bacillus, Pseudarthrobacter, and Streptomyces were dominant, with Bacillus having exceptionally high abundance. These results showed that Cry1Ah1 expression had no significant impact on the quantity of phosphate-solubilizing microorganisms, except for the relative lot of Pseudarthrobacter in lines A5-25, A4-6, and A5-0 (Supplementary Fig. 6). However, soil available phosphorus and MBP content were significantly lower in Cry1Ah1-modified poplars than in NT poplars, possibly because they affected the activity of phosphate-solubilizing microorganisms, reducing the activity of phosphate solubilization and affecting the transformation of soil phosphorus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}