Respective interventions of insulin, glibenclamide and metformin during pregnancy improved the hyperglycemia and insulin resistance in GDM dams.
The whole experimental scheme is shown in Fig. 1A. Based on current data, the growth rate of dams in the untreated diabetic G0 group was significantly higher than those in the drug-treated groups (Fig. 1B), indicating that the respective therapies of insulin, glibenclamide and metformin just exerted substantial reduction effects on the maternal body weights during pregnancy (Fig. 1B). In addition, the fasting plasma glucose levels in untreated G0 group were dramatically higher than those in N0 group, and the interventions of three drugs respectively and dramatically ameliorated this hyperglycemia (Fig. 1C).
As expected, the GDM dams showed the significant hyperinsulinemia, which could be normalized by the respective three drug therapies during pregnancy (P < 0.05, Fig. 1D). Similarly, untreated GDM dams showed significantly elevated HOMA-IR, which could be drastically alleviated by the respective drug therapies (P < 0.001, Fig. 1E).
Respective interventions of three drugs during maternal pregnancy normalized fetal body weights and improved offspring overweight during lactation
After G18, the dams delivered the fetus which were collected and measured respectively. The crown-rump length and body weights of the fetus born to untreated diabetic G group were significantly higher than those in N group (Fig. 2A and B), but the therapies of insulin, glibenclamide and metform in dams respectively normalized these characteristics. Through the lactation (3-21days), the growth rates of offspring born to G group were sharply faster than those in N group (P < 0.05, Fig. 2C), suggesting that the overweight in fetus elicited by untreated GDM in dams exerted the intergenerational impact on their offspring, while the respective interventions of three drugs significantly and similarly ameliorated the effects (P < 0.05, Fig. 2C).
Respective interventions of three drugs during maternal pregnancy normalized the fasting plasma glucose and growth rate in male offspring at adulthood
After three weeks of lactation, male and female offspring were kept in separate cages. Body weights and plasma glucose were measured once a week. The growth rates of female offspring in N, GI, GG and GM groups were gradually reached the same levels including those in the untreated G group (Fig. 3A). Likewise, the fasting plasma glucose levels in female offspring showed the same convergent trend from 4 to 8 weeks of age. These results suggested a gradually autonomous correction of overgrowth and hyperglycemia in female progenies (Fig. 3A and B). However, the male offspring in the untreated G group kept growing faster than those in N, GI, GG and GM groups, and respective interventions during maternal pregnancy showed significant intergenerational improvements on the body weights in male offspring at adulthood (Fig. 3C). Similarly, the male progenies in untreated G group demonstrated the persisted rising hyperglycemia compared to the N group, which could be normalized by the three respective therapies during maternal pregnancy (Fig. 3D).
Respective therapies of three drugs in dams normalized the glucose tolerance and insulin sensitivity in all male offspring at adulthood
In order to examine the offspring’s insulin sensitivity after weaning and when they reach adulthood, we carried out an acute insulin challenge test in the female and male offspring, respectively. As shown in Fig. 4, After exogenous insulin injection in offspring with 4 weeks of age, the plasma glucose levels in the female offspring born to the untreated GDM mice stayed slightly higher than those in N, GI, GG and GM groups (Fig. 4A), while the same tests in male progeny showed significant elevation in untreated group (Fig. 4B). The results suggest that insulin resistance still exist in the male offspring with 4 weeks of age which could be totally corrected by the three respective drug therapies, and that female offspring may exhibit an autonomous alleviation trend. For the same test in the offspring with 8 weeks of age, after the insulin injection, the blood glucose levels in the female offspring born to the untreated G group was not different from those in intervention groups and normal control group (Fig. 4C), indicating an automatic recovery of insulin sensitivity in female adult offspring. While the same test showed that the blood glucose levels in the male offspring born to the untreated dams in G group were still markedly higher than those in N, GI, GG and GM groups, suggesting that the intergenerational intervention effects were still persisting for male adult offspring (Fig. 4D). To explore the offspring’s ability to acutely handle the glucose loading, GTT experiments were performed in mice at 4 and 8 weeks of age, respectively. After exogenous glucose injection into female offspring with 4 weeks of age, the peak of glucose levels in the untreated group appeared at 30 minutes that was slightly different from the 30-minute-peaks in N group and other treated groups (Fig. 4E). On the contrary, the GTT tests in male progenies at 4 weeks old demonstrated substantially higher blood glucose levels in untreated group than N and other treated groups (Fig. 4F). These results suggest that impaired glucose tolerance induced by untreated GDM in dams still exists in the male offspring with 4 weeks of age. Furthermore, the GTT tests conducted in female offspring with 8 weeks of age showed no any significant difference for postprandial glucose levels in all 5 groups (Fig. 4G). Whereas, the same test performed in the male offspring with 8 weeks old revealed that the postprandial glucose levels in GI, GG and GM groups were significantly lower than those in the untreated G group, suggesting that three interventions in maternal mice still elicited the amelioration effects for the male offspring even when they reach adulthood (Fig. 4H).
The normalization of hyperlipidemia and hepatic lipid contents by respective drug therapies in dams during pregnancy still persisted in male offspring at adulthood
The lipid profiles in each group are shown in Fig. 5. For the female offspring with 8 weeks of age, there were no significant differences in serum TG, TC, LDL-C, and HDL-C levels in all groups. These results indicate the possible autonomous correction of dyslipidemia in adult female offspring (P > 0.05, Fig. 5A-D). For the male progenies, compared with N group, serum TG, TC, and LDL-C levels were significantly elevated in those born to the untreated GDM dams (P < 0.05, Fig. 5E-G), while there was no difference in HDL-C levels in each group (P > 0.05, Fig. 5H). For the female offspring with 8 weeks of age, there were no significant differences in liver TG, TC levels in all groups, including those born to the untreated GDM maternal mice (P > 0.05, Fig. 5I and J). For the male progenies, compared with N group, liver TG, TC levels were significantly elevated in the untreated G group (P < 0.05, Fig. 5K and L).
The female adult offspring demonstrated the autonomous normalization of insulin sensitivity, glucose and lipid metabolism
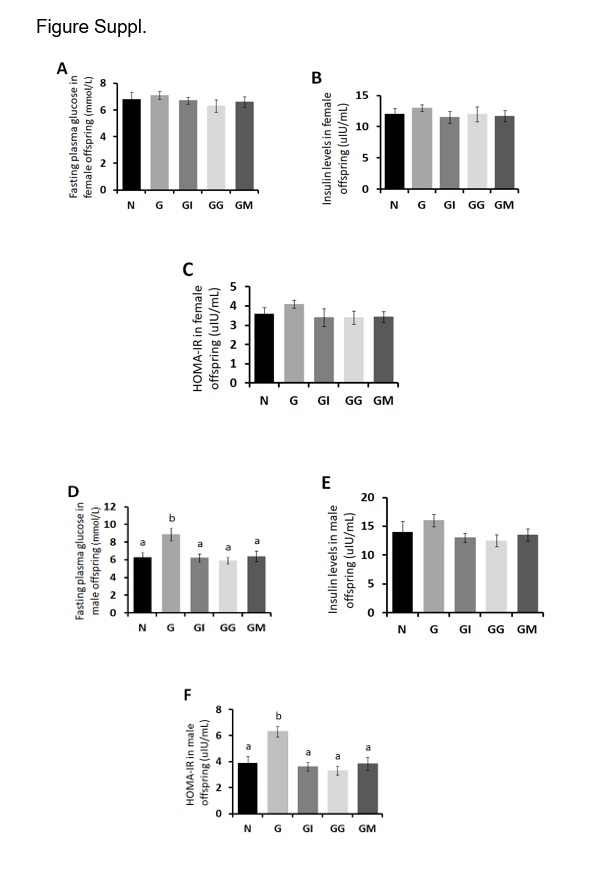
To evaluate insulin resistance in offspring, we measured fasting plasma glucose and serum insulin in male and female progenies with 8 weeks of age. The results showed that there were no differences in fasting plasma glucose and serum insulin levels between each group for all female offspring (P > 0.05, Fig.S1A and B). To explore the mechanism by which the interventions impact insulin sensitivity, we tested the hepatic levels of p-AKT (Thr473, Ser308) and p-FOXO1. Our results showed that the levels of p-AKT and p-FOXO1 in female adult progenies with 8 weeks of age did not differ between all groups, including the untreated G control group (Fig. 6A, and B). PEPCK and G6Pase catalyze committed steps of gluconeogenesis, thus play important roles in glucose homeostasis. The protein expressions of PEPCK and G6Pase in female adult progenies did not differ between all groups either (Fig. 6C). SREBP2 and HMGCS1 play an important role in the regulation of cholesterol synthesis. Our immunoblots showed that the protein expressions of hepatic SREBP2 and HMGCS1 in female adult progenies did not differ between the treated and untreated groups (Fig. 6D). ACL plays a fundamental role in lipogenesis and steroidogenesis. Its activation provides the building blocks for fatty acid biosynthesis. The ratios of p-ACL to ACL did not differ between all groups (Fig. 6E). CPT1A is a rate-limiting enzyme for fatty acid β-oxidation. It can transfer medium- and long-chain fatty acids into mitochondria for β-oxidation, which reduces the deposition of lipids in peripheral tissues. The hepatic protein expressions of CPT1A in female adult progenies did not differ between the untreated and treated groups (Fig. 6F). PPAR- α and PPAR-γ are the nuclear transcription factors. They are not only key regulators of adipose tissue differentiation and lipid metabolism, but also necessary molecules to maintain insulin sensitivity. The hepatic protein expressions of PPAR- α and PPAR-γ in female adult progenies did not differ between the untreated and treated groups (Fig. 6G).
The normalization of insulin sensitivity, glucose and lipid metabolism by respective therapies of three drugs in dams persisted in male offspring at adulthood
The HOMA-IR in female adult offspring from the untreated G group was not different from those in other invention groups (P > 0.05, Fig.SC). However, the male adult offspring born to the untreated GDM maternal mice showed significantly elevated HOMA-IR owing to the substantially enhanced FPG levels in the G group, which could be normalized by the respective therapies of insulin, glibenclamide and metformin during maternal pregnancy (P < 0.05, Fig.SD and SF), despite the similar plasma insulin levels between all groups for male adult offspring (P > 0.05, Fig.SE). Likewise, to explore the mechanism by which the interventions enhanced insulin sensitivity, we found that the hepatic p-AKT (Thr473, Ser308) and p-FOXO1 ratios in male adult progenies born to the untreated GDM dams were still significantly reduced compared with the N group. However, the treatments restored p-AKT and p-FOXO1 levels in male adult offspring with 8 weeks of age, similar to those in N group (Fig. 7A and B). The significant increases in PEPCK and G6Pase were observed in livers of male adult offspring in the untreated G group, compared to those in N group, which were normalized by the three drug therapies during maternal pregnancy (Fig. 7C), except that metformin has no inhibitory effect on the expression of G6Pase in the livers of male offspring (Fig. 7C). SREBP2 in male adult progenies did not differ between all groups (Fig. 7D). HMGCS1 was significantly increased in livers of male adult offspring in untreated G group, compared to N group, and the elevations were sharply inhibited by three drugs respectively (Fig. 7D). The ratios of p-ACL to ACL were significantly decreased in the untreated G group compared to the N group (Fig. 7E), suggesting the increased ACL activity in G group. However, the activity could be dramatically inhibited by the three drug interventions compared to N group (Fig. 7E). The protein expressions of PPAR- α and CPT1A were significantly decreased in livers of male adult offspring in untreated G group, compared to those in N group (Fig. 7F and G). Nevertheless, the PPAR- α and CPT1A levels could be normalized in male adult progenies born to the treated dams, indicating an intergenerational effect by the three drug interventions during maternal pregnancy (Fig. 7F and G). Likewise, the protein expressions of PPAR-γ significantly decreased in the untreated G group compared to the N group (Fig. 7G), which however could be significantly enhanced by the three drug therapies in dams.
{kind=link}