Immunotherapy is a valuable approach for the treatment of cancer. Nanotechnology-based delivery systems emerged as a powerful tool for improving immunotherapeutics. Therefore, their association have been proposed to overcome some biopharmaceutical limitations of immunotherapy. This work aims at designing a novel immunotherapeutic nanoplatform for the treatment of HER2+ breast cancer. Here, purposely-tailored chitosan-shelled nanobubbles (NBs) were developed for the loading of DNA vaccine. The NBs were then functionalized with anti-CD1a antibody to target dendritic cells (DCs). The NB formulations showed sizes of about 300 nm and a good physical stability up to 6 months stored at 4 °C. The in vitro characterization confirmed that these NBs were able to load DNA with a good encapsulation efficiency (82%). The antiCD1a-functionalized NBs targeted to DCs demonstrated the capability to induce activation of DCs both in human and mouse models, and elicit a specific immune response able to delay tumor growth in vivo in mice. The results are the proof of concept that DC-targeted chitosan nanobubbles loaded with tumor vaccine may provide an attractive nanotechnology approach for the future immunotherapeutic treatment of cancer.
Research Article
Exploring Chitosan-Shelled Nanobubbles to Improve HER2+ Immunotherapy Through Dendritic Cells Targeting
https://doi.org/10.21203/rs.3.rs-787377/v1
This work is licensed under a CC BY 4.0 License
You are reading this latest preprint version
Immunotherapy revolutionized the approach to cancer therapy [1]. To date, several immunotherapeutic agents have been approved by the regulatory authorities for the treatment of many types of cancers. Immune checkpoint inhibitors, such as antibodies against cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), Programmed Death-1 (PD-1) and Programmed Death-Ligand 1 (PD-L1) are in clinical use since 2011, and several more molecules, either with inhibitory (e.g., TIM-3, LAG3) or co-stimulatory effect (e.g., OX-40, CD137), are currently in clinical trials [2]. Immune checkpoint modulators dramatically improved the outcome for cancer patients, but they evoke several adverse effects, including immune-related adverse events (irAEs), which often limit their clinical use [3]. More recently, chimeric antigen receptor (CAR) T cell therapies have been approved to treat advanced lymphocytic malignancies [4], and many other are in advance stage of clinical trials also for solid tumor therapy. Despite the remarkable efficacy of this approach for the treatment of leukemia, myeloma and lymphoma, the heterogeneity of tumor microenvironment in solid cancer still remain challenging to overcome for this type of therapeutic agents [5]. The currently available therapies can be improved by targeting other immune cells involved in T cell activation, such as antigen presenting cells (APC) and natural killer (NK) cells [6]. Dendritic cells (DC) are a subcategory of APC particularly interesting for immunotherapy, as they are able to capture and process tumor antigens, allowing for the recognition by T cells and their subsequent expansion to fight cancer [7]. DC-based cancer vaccines have emerged as a potent therapeutic strategy [8], as cancer vaccination offers distinct advantages over standard therapies such as higher specificity, lower toxicity and long-term effects due to immunologic memory [9]. In the development of a DC-targeted vaccine, the selection of the adequate antigen is crucial. Promising results have been achieved both using newly discovered tumor neoantigens [10], and well-established tumor associated antigens [11]. Among the latter, HER2 oncogene is an excellent candidate for the development of vaccines [12, 13], with several formulations already in clinical trials [14]. Furthermore, in order to correctly expand the immune response against tumor, a vaccine needs to effectively reach the DC, which play a critical role in inducing a proper immune activation [9]. Nanoparticle-vaccines might allow to increase antigen delivery to DC and present non-immunogenic nature and sustained antigen releasing ability [15]. Several formulations, including poly(lactic acid) (PLA) / poly(lactic-co-glycolic acid) (PLGA) nanoparticles [16], liposomes [17], and solid lipid nanoparticles [18]. In addition, through decoration of nanoparticle surface with specific antibodies it is possible to convey nanocarriers to DCs, promoting tumor antigen delivery. The liposomal DC-targeting vaccine, L-BLP25, has shown encouraging results in a Phase II clinical trial in patients with advanced non–small cell lung cancer (NSCLC) [19].
Several reports have been made on the use of bubble liposomes (BL) for cancer vaccination [20]. Dendritic cell-based cancer immunotherapy has been proposed as an effective therapeutic strategy for metastatic melanoma and recurrence due to prime tumor-specific cytotoxic T lymphocytes. DCs were exposed to antigens in the presence of perfluoropropane-entrapping liposomes combined with ultrasound and the treated cells were used for prophylactic immunization of mice. Prophylactic immunization with BL/ultrasound-treated DCs provided a four-fold decrease in the frequency of melanoma lung metastases [21].
Complexes with pDNA and mannose-modified BLs have also been utilized for immunization towards cancer of mice in vivo, leading to substantially increased secretion of cytokines TNFα, IFN-γ, IL-4 and IL-6 and also enhanced activity of cytotoxic T lymphocytes [22].
Combined treatments including vaccines and anti-cancer agents have been also studied. The anti-tumor effect of DNA vaccination against melanoma was enhanced using an ultrasound-responsive mannose-modified bubble liposomes in combination with doxorubicin-encapsulated PEGylated liposomes. Effective cytotoxic T-lymphocyte activities stimulated by DNA vaccination were combined with inhibition of tumor growth induced by doxorubicin [23].
Polymer nanobubbles (NBs) are spherical core/shell nanostructures filled by a gas or vaporizable compounds (i.e. perfluorocarbons) with sizes in the nanometer order of magnitude [24]. They have gained an increasing attention for diagnostic imaging and drug delivery, because they can be loaded with drugs, gases and genes for a targeted release [25–28]. Thus, nanobubbles have shown promising results as innovative nanocarriers, with improved loading capability compared to microbubbles, as well as the extravasation capability [29].
Moreover, they might accumulate within tumor tissues through passive targeting, exploiting the Enhanced Permeability and Retention (EPR) effect [29] or through active targeting, binding targeting ligands on NB surface [30].
Indeed, the presence of polymeric shell offers the possibility of shell functionalization with specific targeted ligands.
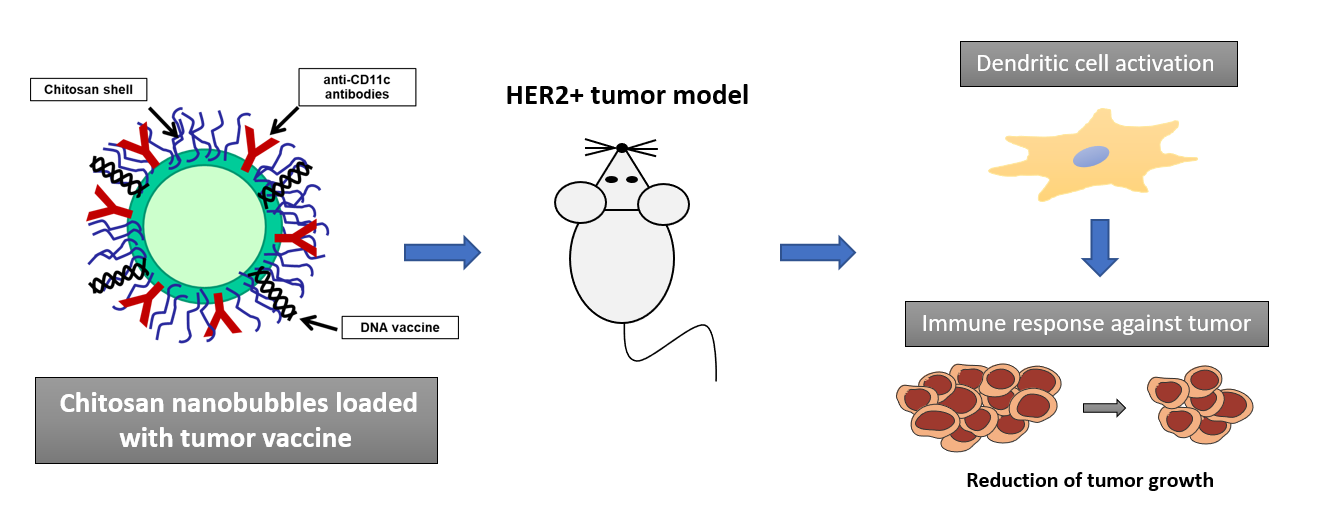
This work aims at designing a novel immunotherapeutic tool exploiting NB technology for the treatment of HER2 + breast cancer. For this purpose, chitosan-shelled NBs loaded with DNA vaccine and functionalized with an antibody to target DCs were developed and characterized in vitro and in vivo.
2.1. Materials
All reagents were of analytical grade and obtained from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise specified. Chitosan medium molecular weight (degree of deacetylation 75–85%, 190–310 KDa) from Sigma-Aldrich was used. Epikuron 200® was kindly provided by Cargill.
2.2. DNA Plasmids
A plasmid coding for GFP (pmaxGFP) and a plasmid pVAX1 coding for the extracellular and transmembrane domains of HER2 as previously described [12] were used for transfection of DCs and in vivo experiments.
2.3. Preparation of nanobubble formulations
NBs were prepared as previously described, using perfluoropentane as inner core component and chitosan for the shell [26]. Briefly, an Epikuron 200 and palmitic acid (1% w/v) ethanol solution was added to perfluoropentane and ultrapure water. The mixture was then homogenized using an Ultra-Turrax® homogenizer (IKA, Konigswinter, Germany). Finally, a 2.7% w/v chitosan solution at pH 5.0 was added dropwise under mild stirring. Interestingly, to obtain the DNA-loaded NBs the pmaxGFP plasmid was incorporated within the chitosan shell of pre-formed NBs by electrostatic interaction. The DNA-loaded chitosan NBs were then functionalized with anti-CD11c or anti-CD1a monoclonal antibody (for in vivo or in vitro experiments, respectively) by amino-reductive method.
2.4. Physico-chemical characterization of nanobubble formulations
Chitosan-shelled NBs, either blank or DNA-loaded (targeted and not targeted), were in vitro characterized. The average diameter and polydispersity index of NB formulations were determined by photon correlation spectroscopy and the zeta potential was measured by electrophoretic mobility using a 90 Plus instrument (Brookhaven, NY). The analyses were carried out at a fixed angle of 90◦ and a temperature of 25°C, after the dilution (1:30 v/v) of the samples with filtered water. For zeta potential determination, the diluted samples were placed in an electrophoretic cell to which a rounded 15 V/cm electric field was applied. The morphology of NB formulations was evaluated by Transmission Electron Microscopy, using a Philips CM10 instrument (Philips, Eindhoven, The Netherlands). Samples were dropped onto a Formvar-coated copper grid and air-dried prior to analysis.
2.5. DNA complexation capacity of nanobubbles
The pDNA complexation with chitosan-shelled NBs was evaluated by gel retardation assay, using electrophoresis in an agarose gel.
DNA-loaded NBs were loaded into agarose gel (1% w/v), stained with an ethidium bromide solution (0.5 µg/mL). The electrophoresis run in TAE buffer (40 mM Tris base, 20 mM acetic acid, and 1 mM EDTA; pH 8.0) at 60 V for one hour. A solution of pDNA (0.1 µg/µL) was used as positive control. The banding pattern was visualized using an ultraviolet transilluminator and photographed with a Polaroid camera.
The amount of pDNA incorporated in NBs was determined spectrophotometrically at 260 nm using an UV-visible spectrophotometer (DU 730, Beckman Coulter, Fullerton, CA). The encapsulation efficiency was calculated by subtracting the amount of free pDNA from the initial amount added, according to the following equation:

The loading capacity was determined on freeze-dried NB samples, according to the equation:

2.6. Determination of NB physical stability over time
The physical stability of blank and DNA-loaded NBs either targeted or untargeted was evaluated over time. The average diameter, Z-potential and morphology of the NB formulations stored at 4°C were determined up to six months.
2.7. Biocompatibility evaluation of nanobubble formulations
To evaluate the hemolytic activity of the NBs, different concentrations of the formulations were incubated with 1 mL of blood diluted with PBS pH 7.4 (1:10 v/v) at 37°C for 90 min. After incubation, the samples were centrifuged (2000 rpm,10 min) to separate the plasma and the amount of hemoglobin released in the supernatant due to hemolysis was measured spectrophotometrically at 543 nm (Du 730 spectrophotometer, Beckman). The hemolytic activity was calculated with reference to complete hemolyzed samples (induced by the addition of Triton X-100 1% w/v to the blood, used as positive control) and negative control (NaCl 0.9% w/v).
Moreover, the cytotoxic activity of NBs was evaluated on HaCat cell line. Cells were cultured at 0.5 × 106 cells/mL in 6-well-plate at 37°C and 5% CO2 in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS), 2 mM glutamine and 1% antibiotics (Gibco, Thermo Fisher Scientific, Waltham, MA, USA). After overnight incubation cells were treated for 24 h with the NBs at different concentrations. The viability of cells was measured by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) assay. The experiments were performed in triplicate and OD was measured at 570 nm using a microplate reader (VICTOR3TM, PerkinElmer, MA, USA).
2.8. Generation and transfection of human dendritic cells
Human peripheral blood mononuclear cells (PBMCs) were isolated by centrifugation over a Ficoll gradient (Histopaque, Sigma) from heparinized venous blood of healthy subjects provided by the local Blood Bank (Torino, Italy). Then monocytes were obtained by MACS magnetic bead separation (Miltenyi Biotec) at a purity of > 93% CD14+. To generate hDCs, monocytes were plated into six-well culture plates (1.5 × 106 cells/mL) (BD Falcon) in RPMI 1640 (Euroclone) supplemented with 10% heat‐inactivated certified FBS (HyClone) and incubated for 5 days, in the presence of GM‐CSF and IL‐4 (PeproTech, both 100 ng/mL). On the day of transfection hDCs were seeded with different amounts of GFP plasmid-loaded nanobubbles targeted or not with CD1a. After 24 hours nanobubbles were removed and medium replaced with fresh RPMI + 10% certified FBS. Transfection efficiency was analyzed at 24 hours by flow cytometry to evaluate the expression of GFP and the viability of DCs. The specificity of CD1a targeted NBs for hDCs, was evaluated by mixing at 1:1 ratio 1 x 106/ml DCs and 1 x 106/ml PBMCs obtained from the same healthy donor in 24 well plates and then transfected them with targeted or not GFP-loaded NBs. Fluorescence was quantitated on a FACSCalibur flow cytometer equipped with CellQuest software (BD‐Biosciences. PBMC and DCs were distinguished according to their physical features. The amount of nanobubbles was evaluated with a NANOSIGHT (“Nanoparticles Tracking Analysis (NTA) Malvern).
2.9. Flow cytometry
To evaluate the ability of GFP-loaded nanobubbles to induce maturation of transfected hDCs, the expression of CD86 and CD83 maturation marker was evaluated by flow cytometry 24 hours after transfection of hDCs with targeted or not GFP-loaded nanobubbles. The following mAbs were used: αCD86-PE (clone IT2.2), αCD83-PE (clone HB15e). Proper isotype-matched control Abs (BioLegend) were used. Cells resuspended with FACS buffer (PBS supplemented with 0.2% BSA, 0.01% NaN3) were incubated with fluorochrome‐conjugated mAbs for 30 min at 4°C, after blocking nonspecific sites with rabbit IgG (Sigma). Fluorescence was quantitated on a FACSCalibur flow cytometer (BD‐Biosciences). Cells were gated according to their light‐scatter properties to exclude cell debris
2.10. In vivo ability of NBs to migrate to lymph nodes
All animal studies were performed in accordance with EU and institutional guidelines approved by the Bioethics Committee for Animal Experimentation of the University of Torino, Torino, Italy and by Italian Ministry of Health (Prot. No. 1009/2015-PR).
BALB/c female mice (Charles River, 6–8 weeks of age) were anesthetized and the back of the animal was shaved to remove the hair and swab with 70% ethanol. Mice received two intradermal injections (one on the left and one on the right side of the back) in the basal tail region 20 µl of GFP-loaded CD11c-NBs or the same volume of naked-NBs as control. At 48 hours three mice per group were sacrificed and inguinal lymph nodes collected. Leukocytes extracted from lymph nodes were stained with anti CD11c-PerCP mAb (Milteniy Biotech) and analyzed by flow cytometry.
2.11. Evaluation of therapeutic efficacy of HER2-loaded NBs
BALB/c female mice were challenged in left flank with a lethal dose (3 x 105) of D2F2/E2 mammary tumor cells expressing human HER2 [31]. Vaccination was started when tumor had reached a mean diameter of 2 mm. Mice received an intradermal injection of 20 µl of CD11c targeted HER2-NBs or empty-NBs or the same volume of PBS, in the basal tail region, two times at 14-day interval. Tumor growth was monitored every 5 days for 30 days after the first vaccination. Two perpendicular tumor diameter was measured with a caliper, and tumor volumes were calculated according to the formula: length x (width)2 x 0,5.
2.12. ELISPOT Assay
Splenocytes (spc) recovered at necropsy of mice vaccinated as above described, were stimulated with D2F2/E2 cells at 10:1 ratio for 48 hours in IFNγ ELISpot assay (BD Bioscience). In order to evaluate CD8-restricted response, D2F2/E2 cells were incubated with MHC-I blocking antibody (Biolegend). Spots were counted with a computer-assisted image analysis system, Transtec 1300 ELISPOT Reader (AMI Bioline). Specific spots were calculated by subtracting spots produced by spc in medium alone from spots produced in presence of tumor cells.
2.13. Statistical analysis
Statistical analyses were performed using Prism 5.0 GraphPad Software. Data are expressed as the median ± SEM. In vivo tumor growth is expressed as mean tumor volume.
In the last decade immunotherapy strategies involving nanotechnology-based approaches to eradicate tumor cells and therefore to improve the therapeutic outcomes have been deeply explored. A number of nanoparticles and nanomaterials have been studied for the targeted delivery of antigens to immune cells increasing the effectiveness of immunotherapy (Mainini et al, 2021, Fogli et al, 2017, Surendang et al, 2018, Zhang et al, 2017). Moreover, the immunotherapy landscape can be furtherly broadened by the combination of delivery systems with external physical stimuli, such as radiofrequency, magnetic fields and ultrasound (US).
In this context, microbubbles have been developed to be associate with US for promoting the localized release and uptake of immunotherapy molecules (i.e. antibody, nucleic acids) promoting immune response [32–34]. Nanobubbles have been designed as a second generation of bubbles being more stable and having the capability to extravasate from blood circulation.
Interestingly, nanobubble technology have been proposed for diagnostic imaging and drug delivery.
Recently, nanobubbles have been investigated to activate immune systems [35–37]. For example, combined multiple anticancer treatments have been achieved using nanobubbles loaded with sonosensitizers and immune checkpoint inhibitors. The antitumor immunity was markedly improved with the NB formulations by maturation of dendritic cells and activation of CD8 + cytotoxic T cells both in vitro and in vivo [35–37]. Here, chitosan-shelled perfluoropentane-cored NB functionalized with anti-CD11c or anti-CD1a monoclonal antibody have been purposely designed for targeted cancer vaccination. This formulation is referred to as “nanobubbles” for sake of simplicity, but it would be more correct to use the term nanodroplets, being perfluoropentane liquid at room temperature (boiling point of 29°C).
Anyway, they can be activated by US that can induce the liquid-to-vapor transition of perfluoropentane by means of a phenomenon called Acoustic Droplet Vaporization (ADV) [38, 39]. The phase-change of perfluoropentane transforms the nanobubbles in a very good reflector for US waves. Therefore, they showed the capability to be visualized by US imaging due to echogenic properties [40].
The main aim of the work was to investigate the capability of this type of targeted nanobubbles to target and transfect dendritic cells.
The average diameter, polydispersity index and zeta potential of the nanobubble formulations, before and after loading with DNA, are reported in Table 1.
The targeted DNA-loaded NBs showed sizes of about 300 nm and a well-defined core-shell structure as demonstrated by TEM analysis (data not shown).
| Formulations | Average diameters ± SD (nm) | PDI | Zeta potential ± SD (mV) |
|---|---|---|---|
| Blank chitosan-shelled NBs | 392.6 ± 17.5 | 0.20 | + 31.90 ± 2.3 |
| pmaxGFP-loaded NBs | 305.3 ± 23.5 | 0.21 | + 15.37 ± 2.3 |
| pmaxGFP-loaded NBs + αCD11c | 311.6 ± 18.7 | 0.19 | + 14.45 ± 1.5 |
| pHER2-loaded NBs | 303.2 ± 21.2 | 0.20 | + 13.68 ± 1.9 |
Interestingly, after incubation of chitosan nanobubbles with plasmid DNA, a marked decrease in size and zeta potential values of about 22.5 and 50 % respectively was observed. These results can be related to the electrostatic interactions between the DNA negative phosphate groups and the positive amino groups of chitosan. This behavior was previously observed with other DNA-loaded NB formulations, either chitosan or DEAE-dextran shelled ones [41]. The polysaccharide multilayered shells showed a marked capability to incorporate and protect DNA embedded in the polymer chains. The presence of DNA condensed the polymer chains, thus confirming plasmid localization and loading in the polymer shell. Here, the pDNA encapsulation efficiency was of about 82 % and the mass of DNA was 3 × 10− 3 µg/µm2.
The pDNA complexation with chitosan-shelled NBs was confirmed by gel retardation assay, using electrophoresis in an agarose gel. The disappearance of the DNA band for DNA-loaded NBs was observed (data not shown). The NB physical stability was confirmed up to six months, by morphological analysis and size and the Z-potential measurements over time. The biocompatibility of the NB formulations was demonstrated by the absence of hemolytic activity and cytotoxicity on HaCat cells.
In order to validate the capacity of DNA-loaded NBs in targeting DCs, we contemporary carried out in vitro in human or in vivo in mice experiments.
At first, we generated human DCs (hDCs) from CD14 + monocytes isolated from venous blood of healthy subjects. Then, hDCs were incubated with different amounts of pmaxGFP-loaded NBs previously conjugated or not with an antibody specific for hDCs marker CD1a. Different amounts of NBs were used, and transfection efficiency was analyzed at 24 hours by flow cytometry. Interestingly, CD1a targeted NBs showed more efficiency in transfecting DCs at all dilution tested, compared to naked NBs (Fig. 1A).
To evaluate the specificity of CD1a targeted NBs to hDCs, we mixed at 1:1 ratio hDCs and peripheral blood mononuclear cells (PBMCs) obtained from the same healthy donor. While naked chitosan-shelled NBs were incorporated likewise by hDCs and PBMCs we observed a preferential transfection efficiency of hDC in comparison to PBMCs by CD1a targeted NBs (Fig. 1B).
Of note, targeted gene delivery exploiting antibody functionalization to direct nanobubbles to specific cells have been previously reported [41–43]. In fact, antibodies conjugated on the nanoformulation surface can enhance the accumulation of nanobubbles in specific tissues.
Definitely, here CD1a-functionalized NBs showed high efficiency in transfecting hDCs with high selectivity for this cell type. This results behavior suggested that NBs can play a crucial role for immunotherapy treatment.
Physiologically, DCs once encounter an antigen (Ag) have to migrate to lymph nodes where they present Ag to Ag-specific T cells and induce T-cell activation and generation. That event requires a deep change in DCs function and phenotypes, also known as maturation. DCs maturation is correlated with up-regulation of cell surface MHC molecules, co-stimulatory receptors and relevant chemokine receptors that improve the ability of DCs to migrate to secondary lymphoid tissue [Patente et al, 2019]. In the absence of maturation stimuli, DCs fail to efficiently elicit T cells response.
Starting from this point, we evaluated the capacity of chitosan-shelled NBs in inducing maturation in hDCs.
We evaluated the expression of costimulatory and maturation markers CD86 and CD83molecules by flow cytometry after transfection of hDCs. Chitosan-shelled NBs can induce an increased expression of maturation marker in comparison to untreated DCs. In addition, the increase is often higher with CD1a-NBs compared to naked-NBs (Fig. 2).
Next, we moved in a in vivo setting. Since murine DCs (muDCs) are characterized by expression of surface marker CD11c, chitosan NBs were decorated with a monoclonal antibody specific for murine CD11c (CD11c-NBs).
Firstly, we assessed the ability of GFP-loaded CD11c-NBs to transfect in vivo muDCs and to elicit migration to draining lymph nodes. Mice were anesthetized and the back of the animal was shaved to remove the hair and swab with 70% ethanol. Mice received two intradermal injections (one on the left and one on the right side of the back) of 20 µl of GFP-loaded CD11c-NBs or the same volume of GFP-loaded naked-NBs as control (Fig. 3A). Mice were sacrificed at 48 hours after injection and inguinal lymph nodes were collected. Leucocytes extracted from lymph nodes were stained with anti-CD11c mAb and analyzed by flow cytometry. The results showed that higher amount of CD11c cells were detected in CD11c-NBs-treated mice compared to control suggesting a migration of muDCs induced by treatment (Fig. 3B). These findings might pave the way to the development of a new strategy enabling the cooperation between nanomedicine and immunotherapy.
To demonstrate the ability of targeted chitosan-shelled NBs in elicit antitumor immune response, BALB/c mice were challenged subcutaneously with D2F2/E2 cells [31], a murine cell line expressing human ErbB-2 (HER2), an oncogene overexpressed in many kind of human tumors [44].
When mice were already displaying established palpable tumors, they received two rounds of intradermal injection of 20 µl of CD11c-NBs loaded with a plasmid coding for HER2 (HER2-NBs) or with an empty vector (empty-NBs) or the same volume of PBS.
Tumor growth were monitored by caliper every 5 days for 30 days, when mice were sacrificed. Mice treated with HER2-NBs displayed a delay in tumor growth compared to untreated mice or mice injected with empty-NBs (Fig. 4).
The specificity of the T-cell response against human HER2 was assessed by IFN-γ ELISPOT assay. Spc recovered at necropsy were stimulated with D2F2/E2 cells. Compared to the control group, spc from mice vaccinated with HER2-NBs secreted higher amount of IFN-γ against HER2 + tumor cells (Fig. 5A) and these cells were mainly CD8 T cells (Fig. 5B).
Chitosan-shelled NBs targeted to DCs have been developed for the treatment of HER2 + breast cancer. They showed the capability to induce activation of DCs both in human and mice, and elicit a specific immune response able to delay tumor growth in vivo in mice. DCs-targeted chitosan nanobubbles loaded with tumor vaccine may provide an attractive nanotechnology approach for the future clinical translation in immunotherapy. This strategy might be of great clinical importance compared to the ex vivo loading of DCs with antigens, that is labor intensive and costly making its application limited. In addition, the visualization by US imaging of the nanobubble distribution might be feasible.
Ethical Statement
Ethical approval and consent to partecipate
All animal studies were performed in accordance with EU and institutional guidelines approved by the Bioethics Committee for Animal Experimentation of the University of Torino, Torino, Italy and by Italian Ministry of Health (Prot. No. 1009/2015-PR).
Consent for publication
Not applicable
Availability of data and materials
The data and materials are available
Competing interests
No competing interest
Funding
This work was supported by the University of Turin funds (Fondi Ricerca Locale ex 60% by RC and MA).
Conflicts of interest
All the authors declare that they have no conflict of interest.
Authors’ contributions
MA and MS formulated and in vitro characterized the nanoformulations, SO, CA, FN performed in vitro cellular and in vivo experiments, MG and RC designed the experiments and gave the intellectual rationale to the work.
Acknowledgements
Not Applicable
- P.N. Kelly, The Cancer Immunotherapy Revolution, Science 359(6382) (2018) 1344.
- R.K. Vaddepally, P. Kharel, R. Pandey, R. Garje, A.B. Chandra, Review of Indications of FDA-Approved Immune Checkpoint Inhibitors per NCCN Guidelines with the Level of Evidence, Cancers 12(3) (2020) 738.
- F. Martins, L. Sofiya, G.P. Sykiotis, F. Lamine, M. Maillard, M. Fraga, K. Shabafrouz, C. Ribi, A. Cairoli, Y. Guex-Crosier, T. Kuntzer, O. Michielin, S. Peters, G. Coukos, F. Spertini, J.A. Thompson, M. Obeid, Adverse effects of immune-checkpoint inhibitors: epidemiology, management and surveillance, Nature Reviews Clinical Oncology 16(9) (2019) 563-580.
- A. Dushenkov, P. Jungsuwadee, Chimeric antigen receptor T-cell therapy: Foundational science and clinical knowledge for pharmacy practice, J Oncol Pharm Pract 25(5) (2019) 1217-1225.
- F. Marofi, R. Motavalli, V.A. Safonov, L. Thangavelu, A.V. Yumashev, M. Alexander, N. Shomali, M.S. Chartrand, Y. Pathak, M. Jarahian, S. Izadi, A. Hassanzadeh, N. Shirafkan, S. Tahmasebi, F.M. Khiavi, CAR T cells in solid tumors: challenges and opportunities, Stem Cell Research & Therapy 12(1) (2021) 81.
- A.D. Waldman, J.M. Fritz, M.J. Lenardo, A guide to cancer immunotherapy: from T cell basic science to clinical practice, Nature Reviews Immunology 20(11) (2020) 651-668.
- R.M. Steinman, J. Banchereau, Taking dendritic cells into medicine, Nature 449(7161) (2007) 419-426.
- M.J. Cannon, M.S. Block, L.C. Morehead, K.L. Knutson, The evolving clinical landscape for dendritic cell vaccines and cancer immunotherapy, Immunotherapy 11(2) (2019) 75-79.
- F. Torres Andón, M.J. Alonso, Nanomedicine and cancer immunotherapy - targeting immunosuppressive cells, Journal of drug targeting 23(7-8) (2015) 656-71.
- M. Peng, Y. Mo, Y. Wang, P. Wu, Y. Zhang, F. Xiong, C. Guo, X. Wu, Y. Li, X. Li, G. Li, W. Xiong, Z. Zeng, Neoantigen vaccine: an emerging tumor immunotherapy, Molecular Cancer 18(1) (2019) 128.
- A. Mougel, M. Terme, C. Tanchot, Therapeutic Cancer Vaccine and Combinations With Antiangiogenic Therapies and Immune Checkpoint Blockade, Frontiers in Immunology 10(467) (2019).
- S. Occhipinti, L. Sponton, S. Rolla, C. Caorsi, A. Novarino, M. Donadio, S. Bustreo, M.A. Satolli, C. Pecchioni, C. Marchini, A. Amici, F. Cavallo, P. Cappello, D. Pierobon, F. Novelli, M. Giovarelli, Chimeric Rat/Human HER2 Efficiently Circumvents HER2 Tolerance in Cancer Patients, Clinical Cancer Research 20(11) (2014) 2910.
- E. Zupančič, C. Curato, J.-S. Kim, E. Yeini, Z. Porat, A.S. Viana, A. Globerson-Levin, T. Waks, Z. Eshhar, J.N. Moreira, R. Satchi-Fainaro, L. Eisenbach, S. Jung, H.F. Florindo, Nanoparticulate vaccine inhibits tumor growth via improved T cell recruitment into melanoma and huHER2 breast cancer, Nanomedicine: Nanotechnology, Biology and Medicine 14(3) (2018) 835-847.
- S. Pallerla, A.U.R.M. Abdul, J. Comeau, S. Jois, Cancer Vaccines, Treatment of the Future: With Emphasis on HER2-Positive Breast Cancer, International journal of molecular sciences 22(2) (2021) 779.
- R. Wen, A.C. Umeano, Y. Kou, J. Xu, A.A. Farooqi, Nanoparticle systems for cancer vaccine, Nanomedicine (Lond) 14(5) (2019) 627-648.
- J. Conniot, A. Scomparin, C. Peres, E. Yeini, S. Pozzi, A.I. Matos, R. Kleiner, L.I.F. Moura, E. Zupančič, A.S. Viana, H. Doron, P.M.P. Gois, N. Erez, S. Jung, R. Satchi-Fainaro, H.F. Florindo, Immunization with mannosylated nanovaccines and inhibition of the immune-suppressing microenvironment sensitizes melanoma to immune checkpoint modulators, Nature nanotechnology 14(9) (2019) 891-901.
- M.A. Oberli, A.M. Reichmuth, J.R. Dorkin, M.J. Mitchell, O.S. Fenton, A. Jaklenec, D.G. Anderson, R. Langer, D. Blankschtein, Lipid Nanoparticle Assisted mRNA Delivery for Potent Cancer Immunotherapy, Nano Lett 17(3) (2017) 1326-1335.
- K. Lee, S.Y. Kim, Y. Seo, M.H. Kim, J. Chang, H. Lee, Adjuvant incorporated lipid nanoparticles for enhanced mRNA-mediated cancer immunotherapy, Biomater Sci 8(4) (2020) 1101-1105.
- R. Sangha, C. Butts, L-BLP25: A Peptide Vaccine Strategy in Non–Small Cell Lung Cancer, Clinical Cancer Research 13(15) (2007) 4652s.
- J. Unga, M. Hashida, Ultrasound induced cancer immunotherapy, Advanced drug delivery reviews 72 (2014) 144-53.
- Y. Oda, R. Suzuki, S. Otake, N. Nishiie, K. Hirata, R. Koshima, T. Nomura, N. Utoguchi, N. Kudo, K. Tachibana, K. Maruyama, Prophylactic immunization with Bubble liposomes and ultrasound-treated dendritic cells provided a four-fold decrease in the frequency of melanoma lung metastasis, Journal of controlled release : official journal of the Controlled Release Society 160(2) (2012) 362-6.
- K. Un, S. Kawakami, R. Suzuki, K. Maruyama, F. Yamashita, M. Hashida, Suppression of Melanoma Growth and Metastasis by DNA Vaccination Using an Ultrasound-Responsive and Mannose-Modified Gene Carrier, Molecular pharmaceutics 8(2) (2011) 543-554.
- M. Yoshida, S. Kawakami, Y. Kono, K. Un, Y. Higuchi, K. Maruyama, F. Yamashita, M. Hashida, Enhancement of the anti-tumor effect of DNA vaccination using an ultrasound-responsive mannose-modified gene carrier in combination with doxorubicin-encapsulated PEGylated liposomes, Int J Pharm 475(1-2) (2014) 401-7.
- R. Cavalli, M. Soster, M. Argenziano, Nanobubbles: a promising efficient tool for therapeutic delivery, Ther Deliv 7(2) (2016) 117-38.
- M.S. Khan, J. Hwang, K. Lee, Y. Choi, Y. Seo, H. Jeon, J.W. Hong, J. Choi, Anti-Tumor Drug-Loaded Oxygen Nanobubbles for the Degradation of HIF-1α and the Upregulation of Reactive Oxygen Species in Tumor Cells, Cancers 11(10) (2019) 1464.
- R. Cavalli, A. Bisazza, M. Trotta, M. Argenziano, A. Civra, M. Donalisio, D. Lembo, New chitosan nanobubbles for ultrasound-mediated gene delivery: preparation and in vitro characterization, Int J Nanomedicine 7 (2012) 3309-18.
- R. Cavalli, A. Bisazza, D. Lembo, Micro- and nanobubbles: a versatile non-viral platform for gene delivery, Int J Pharm 456(2) (2013) 437-45.
- R. Cavalli, M. Argenziano, E. Vigna, P. Giustetto, E. Torres, S. Aime, E. Terreno, Preparation and in vitro characterization of chitosan nanobubbles as theranostic agents, Colloids Surf B Biointerfaces 129 (2015) 39-46.
- F. Marano, M. Argenziano, R. Frairia, A. Adamini, O. Bosco, L. Rinella, N. Fortunati, R. Cavalli, M.G. Catalano, Doxorubicin-Loaded Nanobubbles Combined with Extracorporeal Shock Waves: Basis for a New Drug Delivery Tool in Anaplastic Thyroid Cancer, Thyroid 26(5) (2016) 705-716.
- N. Güvener, L. Appold, F. de Lorenzi, S.K. Golombek, L.Y. Rizzo, T. Lammers, F. Kiessling, Recent advances in ultrasound-based diagnosis and therapy with micro- and nanometer-sized formulations, Methods 130 (2017) 4-13.
- M.P. Piechocki, S.A. Pilon, W.Z. Wei, Complementary antitumor immunity induced by plasmid DNA encoding secreted and cytoplasmic human ErbB-2, J Immunol 167(6) (2001) 3367-74.
- G. Canavese, A. Ancona, L. Racca, M. Canta, B. Dumontel, F. Barbaresco, T. Limongi, V. Cauda, Nanoparticle-assisted ultrasound: A special focus on sonodynamic therapy against cancer, Chemical Engineering Journal 340 (2018) 155-172.
- J. Tu, H. Zhang, J. Yu, C. Liufu, Z. Chen, Ultrasound-mediated microbubble destruction: a new method in cancer immunotherapy, Onco Targets Ther 11 (2018) 5763-5775.
- J.M. Escoffre, R. Deckers, C. Bos, C. Moonen, Bubble-Assisted Ultrasound: Application in Immunotherapy and Vaccination, Adv Exp Med Biol 880 (2016) 243-61.
- W. Um, H. Ko, D.G. You, S. Lim, G. Kwak, M.K. Shim, S. Yang, J. Lee, Y. Song, K. Kim, J.H. Park, Necroptosis-Inducible Polymeric Nanobubbles for Enhanced Cancer Sonoimmunotherapy, Advanced Materials 32(16) (2020) 1907953.
- Y. Tan, S. Yang, Y. Ma, J. Li, Q. Xie, C. Liu, Y. Zhao, Nanobubbles Containing sPD-1 and Ce6 Mediate Combination Immunotherapy and Suppress Hepatocellular Carcinoma in Mice, Int J Nanomedicine 16 (2021) 3241-3254.
- M. Zhao, X. Yang, H. Fu, C. Chen, Y. Zhang, Z. Wu, Y. Duan, Y. Sun, Immune/Hypoxic Tumor Microenvironment Regulation-Enhanced Photodynamic Treatment Realized by pH-Responsive Phase Transition-Targeting Nanobubbles, ACS Applied Materials & Interfaces 13(28) (2021) 32763-32779.
- O.D. Kripfgans, M.L. Fabiilli, P.L. Carson, J.B. Fowlkes, On the acoustic vaporization of micrometer-sized droplets, J Acoust Soc Am 116(1) (2004) 272-81.
- M.T. Burgess, T.M. Porter, Control of Acoustic Cavitation for Efficient Sonoporation with Phase-Shift Nanoemulsions, Ultrasound Med Biol 45(3) (2019) 846-858.
- S. Zullino, M. Argenziano, S. Ansari, R. Ciprian, L. Nasi, F. Albertini, R. Cavalli, C. Guiot, Superparamagnetic Oxygen-Loaded Nanobubbles to Enhance Tumor Oxygenation During Hyperthermia, Frontiers in Pharmacology 10(1001) (2019).
- A. Bisazza, A. Civra, M. Donalisio, D. Lembo, R. Cavalli, The in vitro characterization of dextran-based nanobubbles as possible DNA transfection agents, Soft Matter 7(22) (2011) 10590-10593.
- Z. Song, Y. Ye, Z. Zhang, J. Shen, Z. Hu, Z. Wang, J. Zheng, Noninvasive, targeted gene therapy for acute spinal cord injury using LIFU-mediated BDNF-loaded cationic nanobubble destruction, Biochem Biophys Res Commun 496(3) (2018) 911-920.
- M. Wu, H. Zhao, L. Guo, Y. Wang, J. Song, X. Zhao, C. Li, L. Hao, D. Wang, J. Tang, Ultrasound-mediated nanobubble destruction (UMND) facilitates the delivery of A10-3.2 aptamer targeted and siRNA-loaded cationic nanobubbles for therapy of prostate cancer, Drug Deliv 25(1) (2018) 226-240.
- D.J. Slamon, W. Godolphin, L.A. Jones, J.A. Holt, S.G. Wong, D.E. Keith, W.J. Levin, S.G. Stuart, J. Udove, A. Ullrich, et al., Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer, Science 244(4905) (1989) 707-12.
{kind=link}