The spleen is the largest immune organ in mammals [11] and plays substantial roles in the activation and control of immune responses in the body [12,13]. The spleen can kill a large number of invading pathogens during blood filtration and is the most important organ in the body for defending against Babesia infection [14,15]. Babesia infection causes a series of morphological and physiological changes, such as the enlargement and presence of brown pigment granules [16]. The results in this study showed that the mitochondria in splenocytes exhibited abnormalities after B. microti infection. These results indicated that the spleen suffered very serious damage. With the progression of time, B. microti was gradually cleared by the immune system in the body. We found that the structure of spleen on 19 d of infection gradually recovered to the normal state; for example, the spleen volume gradually decreased, mitochondria also returned to normal, with potential disease amelioration. Although babesiosis is a self-limiting disease [17], damage to the body by B. microti can be substantial from the perspective of degree of spleen injury during peak infection. If hosts have other underlying diseases at this point, host mortality will greatly increase.
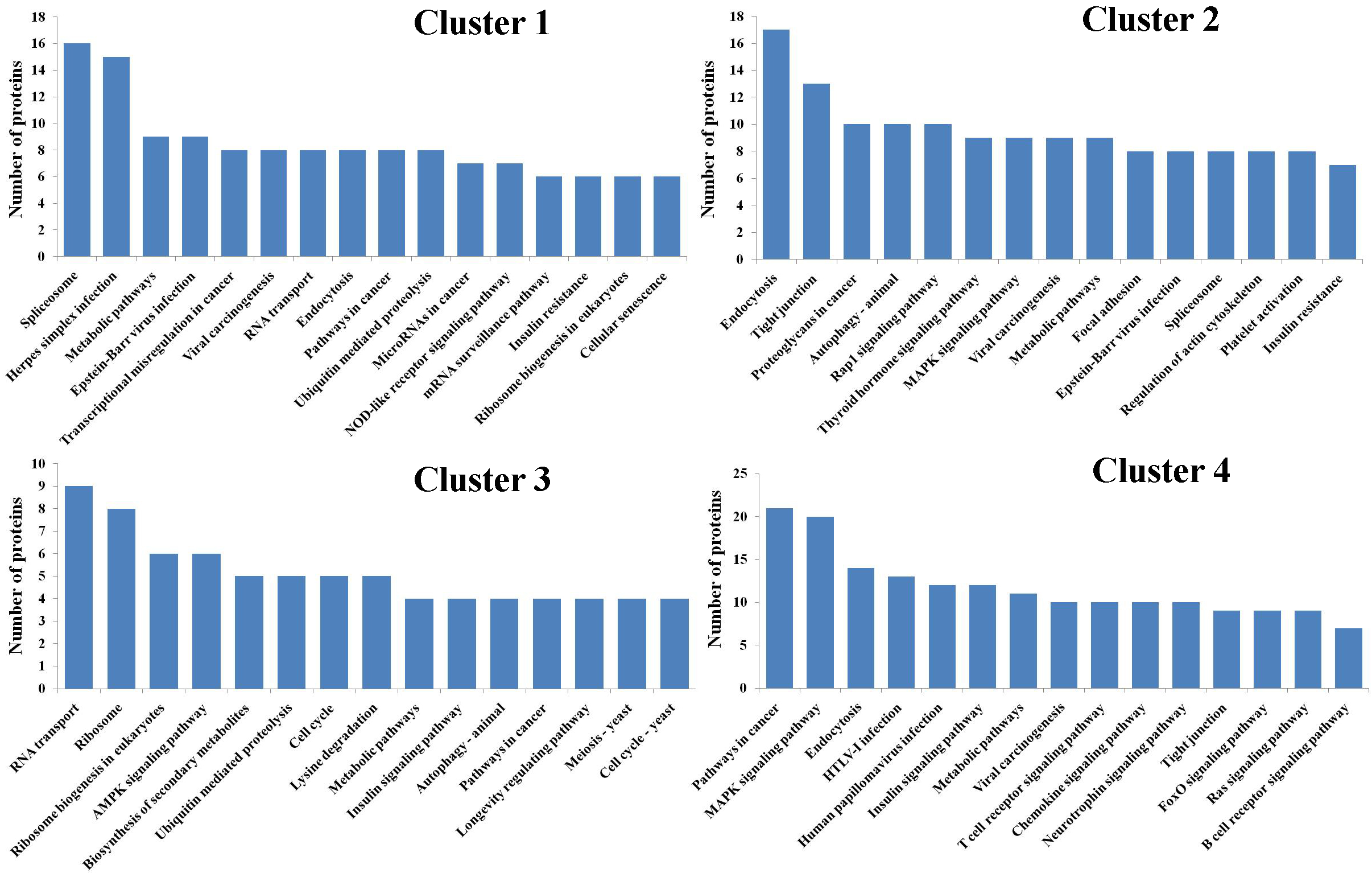
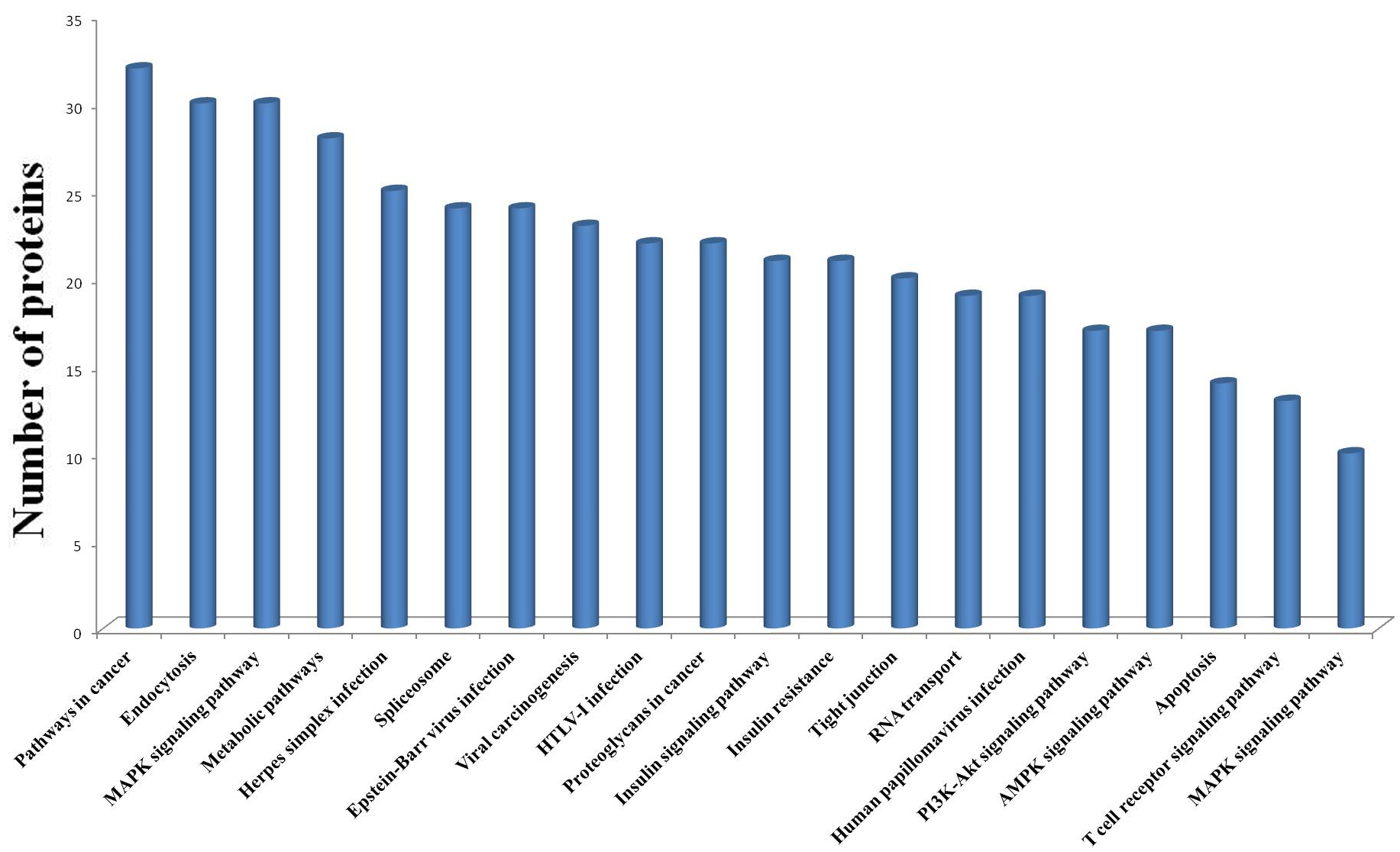
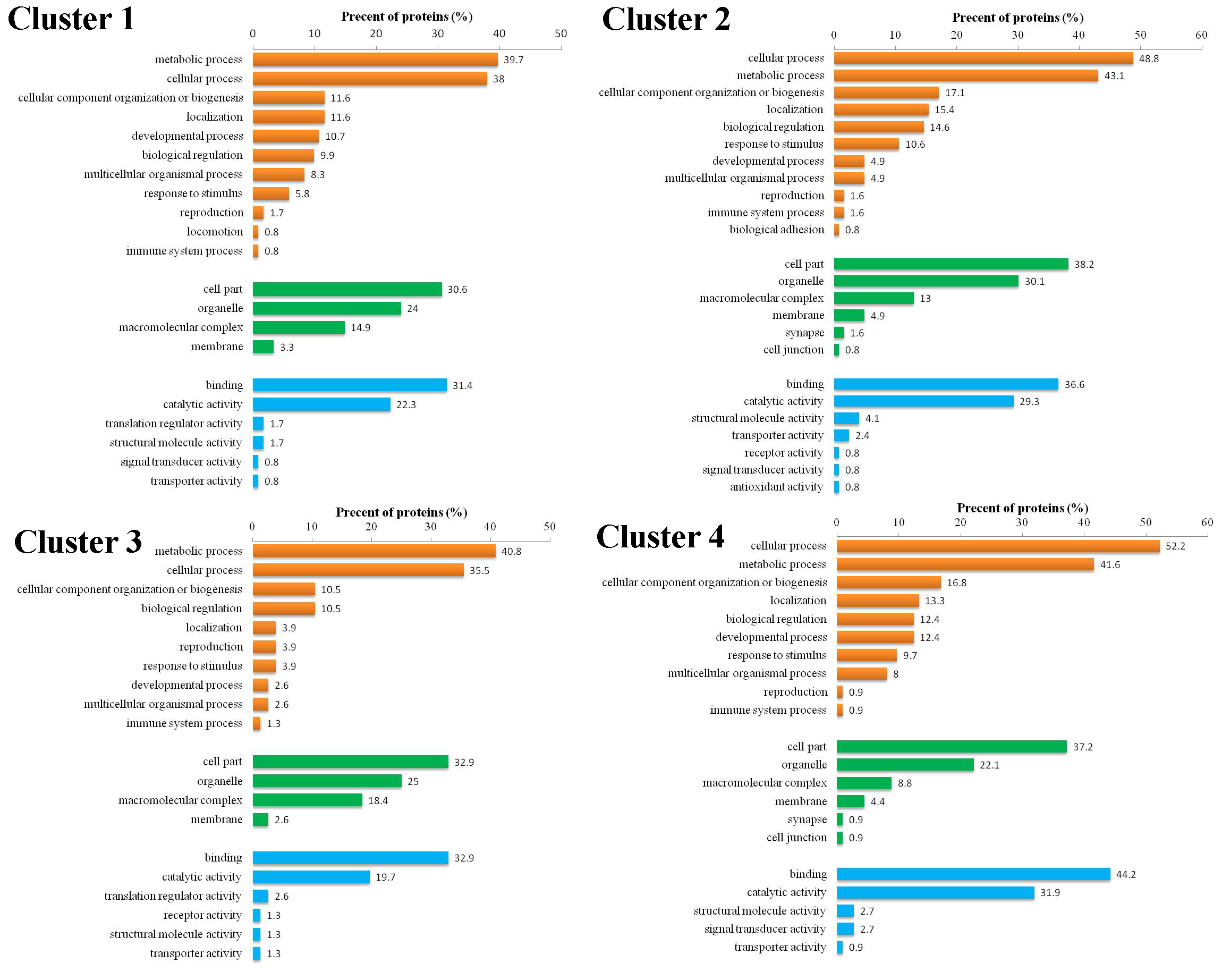
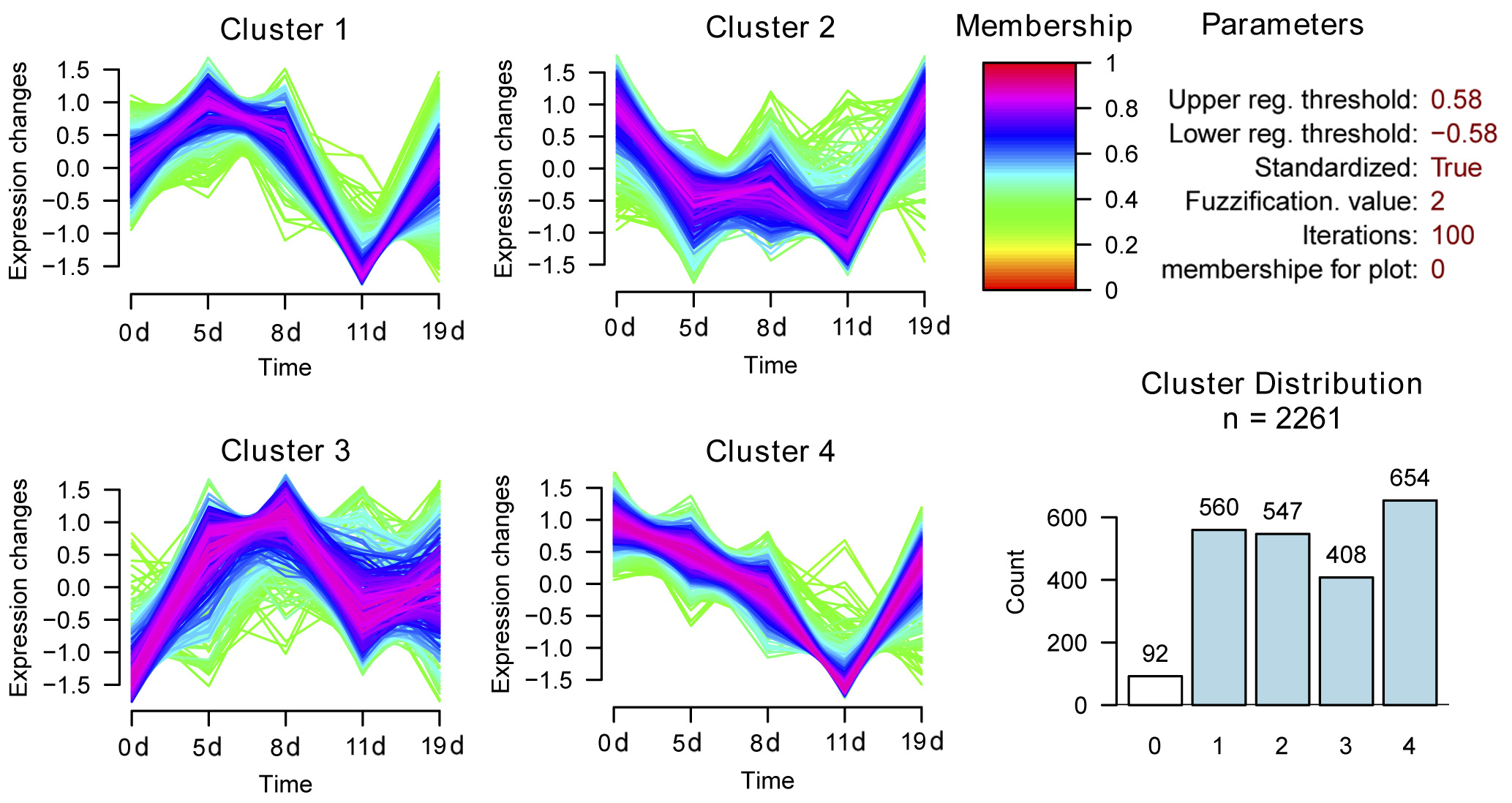
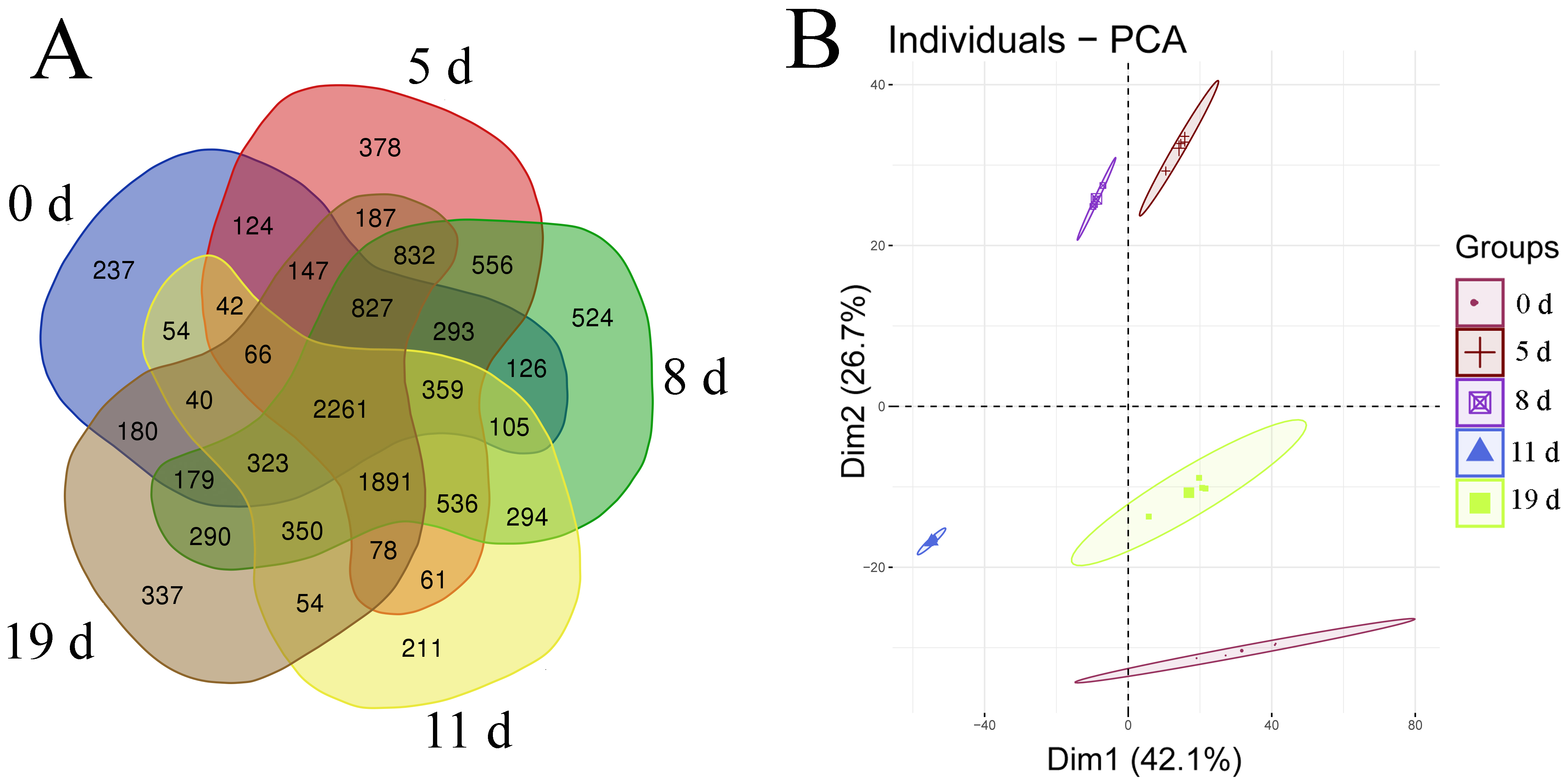
To explore the molecular mechanisms underlying the response of mouse spleen to B. microti infection, DIA quantitative proteomics was performed to systemically study dynamic changes in expression levels of global proteins and protein phosphorylation modifications in spleen tissues of mice after B. microti infection and to investigate protein regulation strategies of the spleen in response to Babesia infection in order to provide molecular bases for more accurate diagnosis and effective treatment of babesiosis. The results showed that the differentially expressed proteins in the spleen after infection were mainly cathepsin, interferon-induced proteins, interleukin enhancing factors, transferrin receptor proteins, glutamate-cysteine ligase, serotransferrin, and growth factor receptor binding proteins. Proteins that exhibited phosphorylation modifications included MAPKs, protein kinases, and transcription factors. These differentially expressed proteins mainly participate in infection defence in the body, the induction of apoptosis and autophagy, the regulation of iron metabolism, and cell proliferation and growth-related pathways.
Immune-related proteins
The quantitative proteomics results in this study showed that the expression of many immune-related proteins changed in mouse spleen during B. microti infection. These proteins included cathepsin D (CTSD), interferon-induced protein 44 (IFI44), interleukin-2 enhancer binding factor 2 (ILF2), interleukin enhancer-binding factor 3 (ILF3), and signal transducer and activator of transcription 5A (STAT5A). These proteins mainly participate in physiological activities such as protein degradation in lysosomes, autophagy, apoptosis, inhibition of excessive cell proliferation, and viral infection defence to ensure effective killing of B. microti.
After hosts are infected with common parasitic protozoans such as Babesia, Plasmodium, and Leishmania, lysosomes in host cells can engulf a large number of parasites [18-20]. CTSD belongs to the aspartic protease family. It is localized in lysosomes of various tissues and cells [21] and can participate in various physiological activities in cells including cell apoptosis [22], autophagy [23], and protein degradation [24]. Some studies have shown that CTSD degrades exogenous pathogens through the autophagy-lysosome system [23]. Furthermore, CTSD can also promote protein degradation in lysosomes to play a critical role in the process of antigen presentation [25]. This study showed that the expression level of CTSD was downregulated on 5 d and 8 d after B.microti infection and were upregulates after 11 d. After the mice were infected with B. microti, the cell structure in spleen was destroyed; in addition, the structure and morphology of organelles such as lysosomes also had abnormalities. We speculate that this might be the cause of the corresponding reduction in the expression level of CTSD in the early stage of infection. With the decrease in the degree of infection, the cell structure in mouse spleen gradually returned to normal. At this time, lysosomes already engulfed a large number of Babesia and a large amount of CTSD was urgently needed for lysosomal degradation of Babesia. Therefore, the expression of level of CTSD was upregulated correspondingly.
IFI44 is an interferonssion oα/β-induced protein [26]. Studies have shown that IFI44 is a potential inflammatory factor and can defend against viral infection through the inhibition of viral transcription [27]. Furthermore, high IFI44 expression promotes the entry of viruses into the latent period and prevents the reactivation of viruses [26]. As an interferon-induced protein, IFI44 expression levels directly reflect IFN-α/β activity. IFN-α/β plays an important role in defending against infection from many parasites such as Leishmania donovani [28], Plasmodium [29], Toxoplasma [30], and Trypanosome brucei [31]. Therefore, it is speculated that IFI44 might have indirect anti-protozoan functions. This study showed that after mice were infected with B. microti, the expression level of IFI44 in the spleen was upregulated on 5 d and 8 d and was then gradually downregulated afterward. The expression level returned to a normal level after 19 d. Therefore, we speculate that IFI44 participated in spleen immune responses after B. microti infection and directly or indirectly exerted its biological functions to defend against B. microti invasion. With the gradual recovery of the body, the expression level of IFI44 gradually returned to normal. Although the mechanism of involvement of IFI44 in defending against B. microti infection is currently still not very clear, IFI44 might be able to be used as a marker for screening Babesia infection.
IFLF2 and ILF3 are components of nuclear factor of activated T cells (NFAT) [32]. It has been shown that the downregulation of ILF2 expression levels inhibits cell proliferation [33], whereas the downregulation of IFL3 expression, in addition to the inhibition of cell proliferation, also inhibits cell migration and invasion and promotes cell apoptosis [34]. Some parasites such as Babesia, Plasmodium, and Leishmania can cause unlimited cell proliferation after infecting hosts to eventually cause diseases in hosts [35,36]. This study showed that after B. microti infection in mice, the expression of ILF2 and IFL3 in the spleen was downregulated; notably, the expression was downregulated to the lowest level on 11 d. The expression level returned to a normal level on 19 d. Therefore, we speculate that B. microti infection in mice might result in excessive cell proliferation; therefore, reduced expression levels of ILF2 and IFL3 were required to inhibit unrestricted cell proliferation and avoid cell lesions.
The quantitative proteomics results for phosphorylated proteins in the spleen showed that phosphorylation modifications of some proteins changed after the spleen was infected by Babesia. These proteins were activated through changes in their phosphorylation modifications; thus, the proteins participated in immune defence. STAT5 is an important transcription factor. STAT5 after phosphorylation modification can initiate target gene transcription to play a critical role in T cell proliferation and differentiation, thus participating in host immunomodulation [37]. The results of this study showed that the phosphorylation modification level of STAT5 in the spleen of mice after B. microti infection was upregulated on 8 d. We speculate that the main function of STAT5 activation was to promote T cell proliferation and differentiation [38] to ensure more T cells participated in immune responses. With the gradual disappearance of Babesia under the clearance function of host immunity, mouse spleen function and structure also gradually returned to normal on 19 d after infection, and the phosphorylation modification level of STAT5 also returned to a normal level. These results indicated that changes in the phosphorylation modification of STAT5 had important immune defence functions during the Babesia infection period, the synthesis of a large amount of proteins was not required, and the normal modification pattern rapidly returned to normal when it was not required in the body. This is a very energy-saving cascade signalling transduction process.
Iron metabolism-related proteins
The iron ion is one of the important trace elements for the maintenance of life activities in the body [39]. Iron deficiency or excess will both cause adverse effects on health [40,41]. It has been shown that iron deficiency affects systemic oxygen transport to cause iron deficiency anaemia to further cause cell death and inhibit body growth [42,43]. Excessive free iron promotes oxygen free radical production, causes lipid peroxidation, oxidative stress, and DNA damage, and eventually leads to cell death, influencing body growth [44,45]. To maintain iron element balance in the body, the body finely regulates iron absorption, transport, utilization, and storage. This series of regulation requires the involvement of many proteins. The major involved proteins include lactoferrin, serotransferrin, transferrin receptor protein 1 (TfR1), and glutamate-cysteine ligase (GCL).
Many parasitic protozoans have the ability to influence iron metabolism in hosts such as Plasmodium and Leishmania. These parasites can uptake a large amount of iron ions in hosts to maintain their growth and propagation [46,47]. When parasites propagate, they utilize hosts to provide enough iron supplements. It has been shown that when the iron content in hosts is too high, protozoan propagation indeed will be promoted [48]. In contrast, when the iron content in hosts is low, the parasitic rate significantly decreases.
Lactoferrin is an important non-haem iron binding protein and participates in the regulation of iron homeostasis in the body [49]. In addition, as an immunomodulatory protein, lactoferrin has many functions, including anti-parasitic [50], anti-bacterial [51], anti-viral [52], and anti-inflammatory [53] actions. Lactoferrin participates in the host defence mechanism through 2 methods. The first method involves lactoferrin binding to iron in hosts so pathogens cannot acquire enough iron from host cells; therefore, their growth is blocked [54]. The other method involves lactoferrin directly interacting with pathogens to inhibit pathogen adsorption and invasion into target cells [52]. The results in this study showed that after B. microti infection in mice, the expression level of lactoferrin in splenocytes was downregulated during the infection period and upregulated during the recovery period. Therefore, we speculate that the reduction in the expression level of lactoferrin in the spleen in the infection period might be caused by the uptake of a large amount of iron in mice by B. microti to supply their growth needs. At this time, iron deficiency in the body caused excessive expression of iron transport-related proteins; therefore, a large amount of lactoferrin was not required. Thus, the expression level of lactoferrin decreased. However, with the extension of infection time, the iron content in hosts decreased dramatically. To maintain iron balance in the spleen during the recovery period, the uptake of iron by cells through various methods is needed. At this time, a large amount of lactoferrin proteins were required to assist in iron transport; therefore, the expression level of lactoferrin was upregulated.
Serotransferrin is a key protein involved in iron metabolism and in defending against microbial invasion in the body [55,56]. It can not only transport iron in the body in a soluble and non-toxic form to participate in iron metabolism [57] but can also inhibit the growth of pathogenic microorganisms through the clearance of free iron ions in hosts [56,58]. The results of this study showed that the expression level of serotransferrin in mouse spleen after B. microti infection was downregulated on 5 d and upregulated on 11 d. This regulation pattern for serotransferrin was similar to that for lactoferrin because they had similar functions in iron metabolism and anti-pathogen activities. They both have iron transport functions [59,60]. In addition, lactoferrin also has iron ion binding functions [50]. The coordinating regulation of these 2 proteins not only inhibited pathogen invasion in the body but also regulated iron balance in the body.
TfR1 distributes on the surface of mammalian cells to mediate the entry of iron taken up by transferrin from outside cells into cells [61]. It has been shown that the expression level of TfR1 is negatively correlated with iron reserve in the body [62]. When iron is deficient in the body, cells increase iron intake through the expression of high levels of TfR1 [43]. In contrast, when iron is excessive, the expression level of TfR1 will decrease correspondingly to decrease iron intake [63]. This study showed that after mice were infected with B. microti, the expression level of TfR1 was significantly upregulated in mouse spleen and the fold upregulation peaked on 11 d. With the gradual recovery of the body on 19 d, the expression level of TfR1 also recovered to a normal level. Therefore, we speculate that the characteristics of expression changes in TfR1 mainly reflect a dramatic reduction in iron reserves due to the consumption of a large amount of iron ions in mouse splenocytes by B. microti after mice were infected with B. microti. To maintain iron ion homeostasis, cells had to express high levels of TfR1 to accelerate intracellular iron ion intake.
GCL is a rate-limiting enzyme of glutathione (GSH) synthesis [64]. It is a heterodimer composed of a modifier subunit (GCLM) and a catalytic subunit (GCLC) [65]. The results of this study showed that the expression of GCLM in mouse spleen on 5 d after B. microti infection was continuously upregulated and returned to a normal level on 19 d, whereas GCLC expression was upregulated on 11 d and returned to a normal level on 19 d. These results indicated that the GCL expression level increased correspondingly after splenocytes were infected with B. microti. With the decrease in the infection level of B. microti, the GCL expression level also returned to a normal level. Infection of hosts by some common parasitic protozoans usually causes abnormal levels of iron in hosts [48], promotes an increase in oxygen free radical levels in the body, and induces oxidative stress responses in the body [66]. GSH protects cells from oxidative damage [67], and the first step of GSH synthesis is catalysed by GCL [68]. Therefore, we speculate that the GCL expression level increased in the spleen after mice were infected with B. microti to effectively accelerate GSH biosynthesis. The presence of a large amount of GSH ensured high oxygen free radical clearance and antioxidant abilities [69] to maintain homeostasis in splenocytes.
Growth and development-related proteins
This study showed that the expression levels or phosphorylation modification levels of many proteins involved in growth and development changed in mouse spleen during B. microti infection. These proteins included protein kinase C-δ (PKC-δ), mitogen-activated protein kinase 3/1 (MAPK3/1), growth factor receptor-bound protein 2 (Grb2), and P21-activated kinase 2 (PAK2).
Protozoan infection usually causes uncontrolled host cell proliferation [70]. When the condition is severe, infection will even result in host organ failure until death. This condition is more common in babesiosis [71]. To prevent excessive cell proliferation, the host body will adopt an effective response mechanism. PKC-δ is a Ser/Thr-specific kinase and is involved in many basic cellular processes, including growth and differentiation [72]. It has been shown that under the function of many cytokines, including IFN-α, PKC-δ is activated by phosphorylation. Activated PKC-δ can inhibit cell proliferation and promote cell apoptosis [73,74]. This study showed that on 8 d and 11 d after B. microti infection in mice, PKC-δ expression in the spleen was not only downregulated but the phosphorylation modification level was also downregulated. We speculate that this pattern of change in PKC-δ during the infection period prevented excessive proliferation of splenocytes during the Babesia infection period to avoid body damage.
MAPK is a protein kinase composed of Ser/Thr kinases [75]. The MAPK signal transduction pathway is linked with cell surface growth factors through Grb2 [76]. MAPK1 (also known as ERK2) regulates cell proliferation, survival, adhesion, and migration through the phosphorylation of hundreds of nuclear substrates and cytoplasmic substrates in cells [77]. MAPK3 (also known as ERK1) also plays a critical role in cell proliferation [78]. We found that the expression level of MAPK1 was slightly downregulated after B. microti infection and that the phosphorylation modification level was significantly downregulated on 11 d. Changes in the expression level of MAPK3 were not large; however, the phosphorylation modification level was significantly downregulated and the fold downregulation was the highest on 11 d. It has been shown that insufficient MAPK3/1 (ERK1/2) expression blocks cell proliferation [78,79]. Dephosphorylation of MAPK3/1 (ERK1/2) inhibits cell proliferation and differentiation [80]. In addition, this study showed that the expression level of Grb2 was downregulated after B. microti infection in mice and returned to a normal level on 19 d. The reduction in the Grb2 expression level reduced the abilities of various cytokines in response to the induction of proliferation signal transduction [81]. Therefore, we speculate that the regulation patterns of these MAPK signal transduction-related proteins in mouse spleen effectively inhibited the unrestricted proliferation of host cells during the infection period.
Babesia infection in host cells results in insufficient blood glucose in hosts [82]. PAK2 is an important participant in the insulin signalling pathway and glucose homeostasis [83]. It has been shown that the downregulation of PAK2 expression promotes glucose uptake [84]. This study showed that after B. microti infection in mice, the expression level of PAK2 in the spleen was downregulated on 8 d and 11 d and was close to a normal level on 19 d. We speculate that the blood glucose level decreased after B. microti infection in mice. Cells reduced PAK2 expression level to promote glucose uptake in order to maintain glucose homeostasis.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}