Relationship between enzymatic activity and discoloration potential
Preliminary tests with four synthetic dyes were carried out in the present work, aiming to find the affinity between T. koningiopsis peroxidase and the structure of the dyes, expecting enzymes to use it as a substrate in the enzymatic bioxidative reaction. Table 2 shows the results.
[Table 2 near here]
The most significant discoloration percentage occurred for the brown dye (54%), with statistical significance, comparing the enzymatic extract amount used (3833 or 7667 U) and the four types of dyes. There was no statistically relevant difference for the red and yellow dyes, but for the blue dye, the enzymatic treatment with 7667 U showed a statistically adequate color removal (22%).
The different discoloration potential observed between dyes indicates the effect of structural diversity. Each dye has a specific molecular structure, which conveys the characteristics of color and dyeing. Considering the molecular aspect, the relationship between the structure and the enzymatic action depends on the type, number, and position (ortho or para) of the groups linked to the aromatic ring of the dyes. In this sense, the molecular aspect can accelerate, delay, or even cause complete inhibition of the enzymatic action26.
Due to its oxidative nature, there are several areas in which peroxidase could replace current techniques based on chemical catalysis27. Although different peroxidases have already been extensively studied, most studies use commercial and high-cost enzymes, making environmental applications unfeasible. In this study, the crude enzymatic extract showed specificity for brown dye discoloration. This result is promising since studies that used peroxidases have reported affinity with other dyes. For example, peroxidase extracted from industrial soybean residues was efficient in degrading blue dye after 3.3 hours of reaction (70% degradation)28, and peroxidase from macrophytes performed well in the degradation of direct azo dyes, such as amaranth and black starch (93% and 87%, respectively, after 120 hours of reaction)29. Also, for the blue dye, two peroxidase sources (Ipomea palmata and Saccharum spontaneum) showed removals of 15% and 70%, respectively, after 1 to 2 hours of treatment, indicating different specificities of plant peroxidase to the same dye30. Enzymatic treatments conducted in other reactional systems also provided different color removal results, as shown by a previous study18, in which rice bran peroxidase removed 39% of the color after 3 hours of reaction in a microwave system, while complete removal was achieved after 24 h in an orbital shaker.
We verified the relationship between peroxidase and laccase activity with color removal in this study. For this purpose, crude extracts with different enzymatic activity profiles were applied to the brown dye discoloration process. In this experiment, the most remarkable discoloration (32%), statistically different from the other assays, occurred with enzymatic activities of 350 U mL-1 and 0.03 U mL-1 for peroxidase and laccase, respectively. Even so, these activities did not represent the most considerable enzymatic amount among the 17 assays performed. The maximum and statistically relevant peroxidase activity was 826 U mL-1, in which the color removal was 7%. For laccase, the maximum value was 0.07 U mL-1, and there was only 8% dye discoloration.
For both peroxidase and laccase, a statistically different enzymatic activity between the assays was observed, which can be explained by the wide range of operational conditions in the fermentation process for the crude extracts production, such as temperature, pH, and agitation. Thus, the fermentation manipulation can provide an enzymatic pool with different performances in application processes, although the maximum activity did not represent the maximum discoloration potential.
Similar findings were observed when manganese peroxidase (MnP) and manganese independent (MIP) activities were evaluated31. The authors found a difference between the enzymatic activity and the discoloration of synthetic sulfonephthalein dyes. The variations in the discoloration reflect the differences in the isoenzyme composition of the MnP and MIP, resulting in a difference in the kinetic constants (varying the maximum rate of reaction from 0.057 to 1 U mL-1) and the substrate specificity31. Another study also concluded that the reaction conducted with laccase alone did not degrade certain types of dyes. When the redox mediator violuric acid was added, however, the degradation efficiency reached 90% using 2/5 of the enzymatic activity of the tests without mediator32.
Our results thus indicate that the change in operational parameters in fungal fermentation, in addition to providing enzymatic activity variations, must have influenced the composition of isoenzymes. In turn, it caused differences in specificity and interfered with the decolorization activity. It is noteworthy that until this step of the work, no enzymatic manipulation method had been employed. Later on, the reactions with immobilized peroxidase on different supports will be discussed.
Catalytic properties of free and crude peroxidase
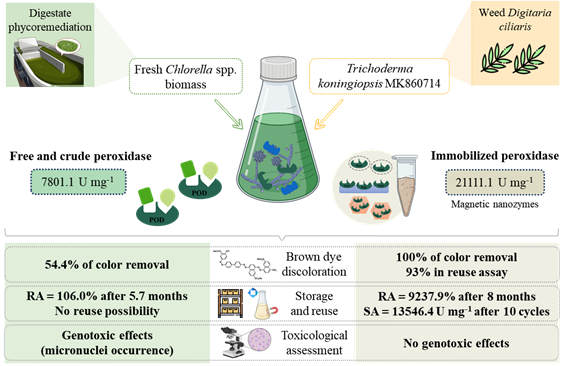
To insert a novel enzyme source in biotechnological processes, it is essential to know and to improve its catalytic properties and reactional conditions. This work is the first to present the preliminary characterization of guaiacol peroxidase produced by T. koningiopsis in submerged fermentation supplemented exclusively by fresh microalgal biomass.
The extracellular enzymes of this fungus are rarely mentioned in the literature due to the difficulty in offering favorable conditions for fermentation and supplementation for its expression, often involving costly processes for industrial applications5. Mäkelä et al.33 report a wide variety of fungal peroxidases being applied as biocatalysts to reactions of environmental interest. In their study, the species Trichoderma is classified as Soft-Rot Fungi, about which little is known concerning peroxidase production. Azmi et al.34 reported a combination of lignin peroxidase production in a more significant proportion and manganese peroxidase in a smaller proportion by T. koningiopsis when supplemented with oil palm fronds in solid-state fermentation.
Regardless of the peroxidase source, enzymatic processes are influenced by reaction thermodynamics and need studies varying pH, temperature, and availability of substrate and cosubstrate - relevant factors in understanding the catalytic route35. In this sense, these variables were investigated in the present work for the T. koningiopsis POD following a Plackett-Burman design. The results indicated enzyme stability even when subjected to adverse reaction conditions.
When subjected to pH ranging from 4.0-8.0 and temperatures from 20-80 °C, the enzymatic activity was not significantly affected at 95% confidence level by the Tukey test. The enzyme:substrate and enzyme:cosubstrate ratios, conversely, negatively affected the enzymatic response with statistical relevance; that is, the POD activity was improved at the minimum levels studied. None of the reaction conditions led to enzyme denaturation, with the lowest activity obtained being 5733 U mg-1 in a test with pH 4.0, temperature 20 °C, and maximum levels of substrate and cosubstrate.
The peroxidase stability under varying conditions can be explained by the defense mechanisms previously developed by the fungus in its natural habitat36. Our enzyme was isolated from weeds already adapted to its ecosystem. The supplementation of the fermentation medium with microalgae biomass induced higher peroxidase productivity compared to synthetic supplementation37, confirming that stress induction during the fungus growth improves peroxidase production.
In contrast to the production behavior of other enzymes, stressful environments induce higher productivity of oxidoreductase enzymes and, consequently, fermentation conditions with less nutrient availability can result in greater peroxidase expression. As well as the fungus fermentation process, the microalgae growth process occurred in a stressful environment, considering that it was cultivated with mainly ammonia and phosphorus availability13. Thus, according to a previous study14, synergism between fungi and microalgae occurs, resulting in microalgae cells incorporated into the fungal hyphae mesh. This synergy may have led to the obtention of an enzymatic extract with high resilience capacity in peroxidase activity, which can be seen in Fig. 1, where the interaction between the enzyme and reaction medium over time is shown under constant environmental conditions.
[Fig. 1 near here]
To ensure that the evaporation or natural oxidation of the enzymatic extract did not induce false positives, controls were performed and considered in the enzymatic activity quantification. During the follow-up of the reaction, in the first 192 h, the pH and ORP values were maintained without statistically significant differences. The pH remained between 7.3 ± 0.1 and 7.6 ± 0.1, and the ORP between 195.6 ± 3.3 and 214.2 ± 1.1 mV vs. SHE. In turn, the enzymatic activity varied from 7801 ± 179 to 11 ± 2 U mg-1, significantly decreasing and reaching its minimum value in 96 h.
In the last experimental point studied (360 h), the pH significantly dropped to 6.5 ± 0.4, and the redox potential increased to 262.0 ± 20.4 mV vs. SHE. For this experimental point, the standard error became high and, therefore, the experiment was stopped. In this sense, it is understood that after 360 h, it is unfeasible to maintain the enzymatic reaction without providing maintenance.
The constant decrease in enzymatic activity in the first hours of reaction follows the same behavior as observed in a published study for guaiacol peroxidase extracted from rice bran38, in which in 24 h of reaction, the activity had already fallen more than half of the initial value. In the present study, intra and extracellular enzymatic system of the fungus can be critical in conducting the catalytic route through the quinone redox cycling mechanism. The presence of lignin and Fe2+ in class II heme peroxidases structure induces a reaction that produces hydroxyl radicals in the presence of hydrogen peroxide. The enzymatic behavior shown in Fig. 1 can thus configure an advanced bioxidative process, in which the cosubstrate presence generates hydroxyl radicals, which in turn can regenerate cosubstrate. Thus, the enzymatic reaction is naturally maintained, without the need of external interventions39.
The maintenance of positive ORP values during the reaction suggests that the reaction medium was receiving electrons, and the substrate was oxidized. At the end of the reaction, the increase in ORP may have been a reflection of pH decrease, these parameters being inversely proportional40. The change in these parameters may have triggered the enzymatic reaction instability, which had already carried out multiple quinone redox cycles.
Considering the promising results of peroxidase activity and reaction maintenance even in adverse conditions, studies were conducted to monitor the enzymatic activity of the crude extract after storage. The results of this characterization are shown in Fig. 2.
[Fig. 2 near here]
The enzymatic extract activity (initially 7801 U mg-1) remained more stable at room temperature and in the refrigerator than in the freezer. For the storage condition between 18-28 °C, there was no significant difference in activity up to the 60th day, reaching 105 ± 3% of residual activity. For storage at 4 °C, a considerable drop was observed between 30 and 90 days (91 ± 1%), but on the 170th day, the RA returned to the initial value. In turn, the extract maintained at −10 °C, initiated a significant and current drop of enzymatic activity after 15 days, reaching 63 ± 1% of RA in 90 days.
From these tests, it was possible to determine that the enzymatic extract can be kept in the refrigerator for up to 170 days without sudden drops in peroxidase activity. In addition, keeping the extract at room temperature proved to be an interesting option for up to 60 days, but it still requires studies for longer periods, as on the 170th day, a significant drop was observed. Therefore, the activity behavior after storage will be discussed in the next section.
Strategies for T. koningiopsis peroxidase immobilization
Three different strategies were selected to conduct the immobilization process of crude peroxidase extract (Free-POD), obtained in the fermentation process of Trichoderma supplemented by Chlorella spp. Two parameters were chosen to determine the immobilization efficiency: the residual activity and the discoloration potential of the brown dye solution at 100 mg L-1. This treatment process was selected because of its promising results in crude extract assays, the scarcity of studies on this dye, and the objective of inserting this bioproduct in the context of low-biodegradability contaminant remediation.
The immobilization strategies of this study were selected, taking into account the cost-benefit in the scenario of effluent treatment and circular economy. The strategies allowed obtaining relevant results, with an increase in specific activity of up to 589% and color removal of up to 100%, as shown in Table 3.
[Table 3 near here]
RA values up to 100% indicate enzymatic retention in the support for the technique studied, and higher values demonstrate a positive interaction between support and enzyme. For the encapsulation method, it was possible to retain only 6% of enzyme in the POD-beads, and 22% was lost to the calcium chloride solution. When the technique was modified (ModPOD-beads), adding the POD substrate and cosubstrate to the beads to manipulate the affinity between enzyme and support, the retention was lower (3%), as well as the activity in the solution (21%). These results indicate that the encapsulation technique is not suitable for Trichoderma peroxidase because of the low enzymatic retention in the beads. There was also a loss of enzymatic activity in the process, since the RA balance considering beads and solution does not reach the reference value (100%).
Two different supports were studied in the adsorption method: the commercial clay MK10, widely used in enzymatic immobilization studies, and noncommercial ornamental rock dust (OR). The choice of this alternative support was motivated by the environmental problem related to the disposal of this waste material, which is constantly generated by rock processing industries. For both supports, enzymatic activity was lost to the buffer solution, in which the contact process with peroxidase was conducted (19% for MK10 and 15% for OR). However, the commercial support had a greater affinity with the enzyme, with a 35% activity retention, when compared to rock dust (11%). When making the RA balance, a loss of activity was observed in this case. However, compared to the encapsulation strategy, adsorption proved to be more promising and retained higher enzymatic activities in the supports.
The most advantageous process from the perspective of maintenance and even of increasing enzymatic activity after immobilization was by magnetic nanozymes synthesis, conducted with low-cost reagents. In this strategy, although part of the activity was lost to the solution in which the nanoparticles precipitated, the bond between POD and support favored the expression of catalytic activity. This process made it possible to obtain 21111 U mg-1 of specific activity in MN-POD, representing a RA of 689%, and relevant potential for application in biotechnological processes due to their magnetic properties.
As in the present study, commercial horseradish peroxidase was immobilized by magnetic nanocomposites by Chang et al.41, exhibiting biochemical properties superior to those of the free enzyme, maintaining the RA in 100% and improving resistance to temperature and pH variations. In turn, Monteiro et al.42 investigated the immobilization of commercial Candida antarctica lipase onto magnetic nanoparticles and achieved RA values of up to 120% for a pH range between 8 and 10. This same RA value was achieved for POD magnetic nanozymes in Sadaf et al.21 study, however for a pH range between 3 and 4. Although for each nanoparticle synthesis study, different methods are used, and will give them particular biochemical characteristics, Zdarta et al.43 highlight that the presence of many functional groups in the materials used for the synthesis gives to the nanozymes a high surface area and porosity. These characteristics may justify the fixation of a large amount of enzyme in the nanoparticles and the increase in catalytic activity compared to the free enzyme.
Among the techniques studied in the present work, MN-POD showed the most significant potential for discoloration, removing 100% of the color of the dye solution in 5 hours. However, in this enzymatic treatment, the acidic pH of the post-treatment solution would require adjustment before its final disposal. For treatments with POD immobilized by encapsulation and adsorption techniques, the pH remained close to neutrality; however, color removals of only up to 42% were obtained.
From the data presented in Table 3, it is not possible to determine a direct relationship between RA and DP values. It is worth mentioning that blank control tests with dye solution and support, in the absence of enzyme, were conducted. However, they did not bring conclusive contributions, since the blank control results were mostly influenced by pore size and contact surface. The results of the enzymatic treatment depend on the cross-interaction between support, enzyme and dye, giving complexity to the process to be investigated in future works. Accordingly, in addition to enzymatic retention, the immobilization process becomes relevant to provide a combined action between support and substrate that enhances the catalytic reaction. In addition to the enzymatic activity retention, the immobilization process must provide the possibility of reusing and storing the enzyme, maintaining catalytic efficiency. In this sense, MN-POD was applied to the second batch brown dye treatment, in which 94% of discoloration was obtained in 5 hours, maintaining the same conditions as the first batch.
In the storage test, MK10-POD, OR-POD, and MN-POD samples were stored at room temperature protected from direct sunlight, and their activity was measured after 2.5 months of the immobilization process. Comparing to free-POD, the MK10-POD, OR-POD, and MN-POD remained with residual activities of 193%, 3%, and 10637%, respectively, after the storage period. Additionally, MN-POD activity was verified after about 8 months, and it has 9238% of RA.
The data in Table 3 reveal that OR-POD lost enzymatic activity after storage, while MK10-POD and MN-POD increased it. For all samples, the specific activity of the immobilized enzyme remained higher than in free form. The same behavior of activity increase after storage was observed for the crude extract. This phenomenon can be attributed to changes in the biochemical conformation of the protein or to an increase in specificity with the substrate, which is already reported in the literature for enzymes submitted to upstream processes, such as concentration and enzymatic purification8.
The MN-POD sample stored for about 8 months, with 283256 U mg-1, was subjected to 10 cycles of recovery and reinsertion in a reaction medium to monitor the residual activity behavior. In cycles 2 and 3, a statistically significant decay in RA was observed, arriving at the end of cycle 3 with 37%. From cycles 4 to 10, there was no considerable decay in RA. At the end of the experiment, the MN-POD had 13546 U mg-1 of specific activity, corresponding to 5% of RA. Both activity and degradation efficiency decay are expected in reusability assays, considering that the enzyme active site and the support pore voids are filled over cycles9,44.
In this study, the enzymatic activity improvement was possible using simple immobilization techniques and interesting cost-benefit, factors of great relevance in industrial applications. In addition, the bioproduct storage under mild conditions, dispensing chemical supplement addition, or freeze-drying processes, are advantages to its insertion in the enzyme market for industrial applications, which is in constant expansion according to the latest Global Industry Analysts report4.
Support and immobilized peroxidase characterization
OR-POD, MN-POD, and MK10-POD samples were submitted to characterization to understand the interactions between enzyme and support. In addition, support samples without enzyme were also characterized.
The FTIR and XRD graphs are presented in Fig. 3 and, from a first examination, it is possible to observe the similarities when comparing the spectrum of the support and support with enzyme; the transmittance peaks are in the same wavenumber range in FTIR analysis, as well as the intensity peaks in the same 2q values in XRD patterns.
These findings indicate that the enzyme-support interaction did not modify the characteristic chemical bonds or the structural arrangement of the support material atoms during the immobilization processes. This aspect is positively relevant considering that immobilization aims to maintain both enzymatic and material properties. In the current work, this objective was achieved, and the immobilization maintained peroxidase catalytic activity without affecting the physical-chemical characteristics of the materials.
[Fig. 3 near here]
Another relevant observation is the similarities, for the two analyses, of the peak behavior for the adsorption methods (samples OR-POD, OR, MK10-POD, and MK10), indicating that ornamental rock dust is very similar to the commercial and widely used clay MK10. The FTIR spectra of these samples show weak peaks between 820 and 947 cm-1 (Fig. 3a), corresponding to Al−Al−OH, Al−Fe−OH and Al−Mg−OH deformation, and Si−O stretching mode. The absence of peaks around 1000 cm-1 indicates no water content in the samples45.
For MN and MN-POD samples, small intensity peaks were observed at 935, 1059, and 1144 cm-1, suggesting the presence of a significant amount of specifically adsorbed sulfate groups. These can occupy external and internal surfaces (sulfated goethite), and the amount of these groups is related to the acidic pH. The peak at 1591 cm-1 indicates H2O bending vibrations46.
In XRD analysis of magnetic nanoparticles, the peaks indicated in Fig. 3b, between 27 and 50°, confirm goethite presence as a single phase, characteristic of superparamagnetic particles47. Also, characteristic peaks corresponding to planes of Fe3O4 and MgO crystals were observed, whose low intensity is due to the small size of the synthesized particles48.
The amorphous characteristic of the MN and MN-POD samples is another positive relevant finding from the XRD analysis. It is beneficial for many applications due to the superior catalytic activity, superparamagnetic behavior, and large surface area compared with crystalline structures49. Experimental conditions are relevant in amorphous Fe3O4 synthesis, especially regarding particle size and structure homogeneity50. In this sense, the method we used can be considered adequate for obtaining excellent magnetic nanoparticles.
From the XRD patterns in Fig. 3b, it is possible to verify very similar crystalline structures of OR-POD, OR, MK10-POD, and MK10. The 2q values variation between samples does not exceed 0.5 degrees, but there are some differences in intensity. The main highlight is the occurrence of an intense peak at 60.2° for OR, confirming that this material has SiO2, as expected, since it is the primary mineral of this material. Small intense peaks associated with SiO2 are also clearly observed at 26.7 and 27.5°, both for OR and MK10. The remaining low intense peaks in Fig. 3b are natural mineral signatures and were expected for these materials45.
Toxicological assessment of enzymatic treatments
The test organism selected was A. cepa, in which disorders in cell division can be counted and compared using the mitotic index. With a negative control as a standard (cells exposed to tap water), higher or lower mitotic index (MI) values indicate disorders in the mitosis process, as shown in Table 4.
According to Fiskesjö22, one of the precursors of this test, the meristematic onion cells study, is indicated for environmental monitoring, presenting advantages such as low cost, easy execution, and possibilities of microscopic research, being possible to evaluate the chromosomal damage caused by an aqueous matrix and disorders in cell division.
[Table 4 near here]
By qualitative observations on slides, it was possible to detect chromosomal aberrations caused by enzymatic treatments. In Fig. 4, these damages are indicated to emphasize them in some of the treatments. The presence of micronuclei in free-POD treatment is one of the main genotoxic highlights, indicating that the crude enzymatic extract can induce uncontrolled cell division and possibly tumor formation51. In contrast, chromosomal aberrations were not observed for the other treatments, indicating that enzymatic immobilization is relevant to eliminate genotoxic effects. However, for all assays, disorders in the mitosis process were quantified, as shown in Table 4.
[Fig. 4 near here]
In the MN-POD assay, for which complete discoloration was achieved, the acceleration phenomenon of cell division was quantified, increasing by 5.0 times compared with the negative control. When the sample was diluted at 1:8, a delay in cell division occurred (0.2 times), being possible to verify this by MI values (Table 4) and visual comparison between images (a), (c), and (d) of Fig. 4. As in negative control (tap water), for the enzymatic-treated samples, the cells were predominantly in the prophase phase, i.e, the first stage of cell division. In turn, in the dye solution before treatment, all the division phases were visualized, although the MI value was only 1.1 times greater than the negative control.
The findings regarding the cell division phases are relevant since cells in metaphase and anaphase are more susceptible to chromosomal changes and DNA abnormalities52. In this sense, cytotoxic and genotoxic analyses provide complementary results since mitotic indexes close to the negative control value do not guarantee the absence of chromosomal aberrations. According to Fiskesjö22, toxicological assessment can provide a diagnosis of possible behaviors of the test organism when exposed to treatments, but it is a result of multiple factors combined. For example, the author found a relationship between the pH of the solution and toxicological effects; however, it was not possible to find this direct relationship between disorder in cell division and pH in our work.
Miranda-Mandujano et al.28 and Feng et al.53 attributed the increase in toxic effects after enzymatic reaction to the formation of transformation products, during the catalytic route. It is believed that in our study, these products with distinct toxic effects may have been formed after enzymatic treatment because of the disorders caused by cell division. These disorders are possibly related to the different enzymatic conformations obtained by the immobilization strategies, considering that other cytotoxic behaviors occurred between tests, even with dye concentration and operating conditions being equal.
The samples dilution before exposure to A. cepa cells resulted in a change in cell division behavior, as indicated by the results in Table 4. It is understood that the search for a dilution factor in the context of the treatment proposed in this work can be one of the strategies for reaching MI values closer to the negative control. It is worth mentioning that the degradation of different compounds by the enzymatic route presents different responses from the toxicological point of view. Therefore, it is relevant that treatments inserted in other contexts are deeply analyzed considering multiple aspects.



{kind=link}