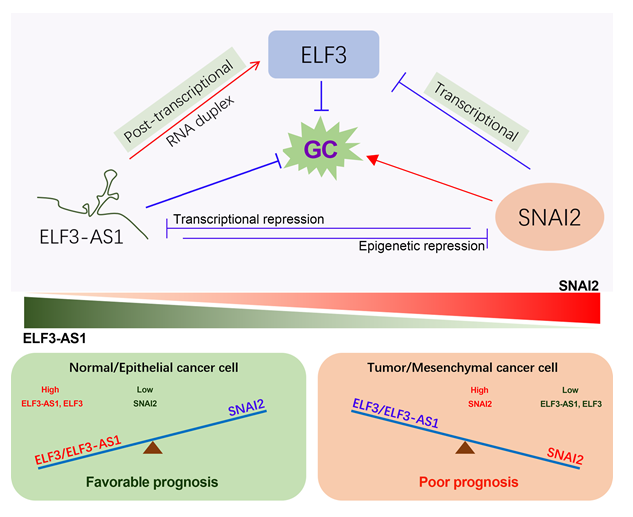
Exploration of lncRNAs regulated by transcription repressor SNAI2 in GC
To explore the lncRNAs regulated by SNAI2, RNA sequencing studies were conducted in the GC cell line overexpressing SNAI2. A total of 318 coding genes, 70 lncRNAs were strongly repressed by SNAI2, while 55 coding genes and 53 lncRNAs were greatly upregulated by SNAI2 (|Log2FC| >1, Table S3). As shown in the Fig. 1A, the expression of IGFBP1/3, ITGB4, CLDN1/4, TJP3, EFNA1, KRT80, ELF3 etc. were strongly repressed by SNAI2, while the expression of CYR61, CTGF, IL11, ID2/3, RBM3, THBS1 etc. were strongly upregulated by SNAI2. Similarly, around 123 lncRNAs, including linc01315, MIR210HG, FZD10-DT, linc00963, HOXB-AS3, LUCAT1, ITPR1-DT, PDCD4-AS1, ZNF197-AS1 etc., greatly altered their expression after overexpressing SNAI2 (|Log2FC| >1). Among them, ELF3-AS1 was one of the top 10 lncRNAs that strongly repressed by SNAI2 (Fig. 1B).
The biological function of lncRNA ELF3-AS1 remains largely unknown in cancers. To further confirm whether ELF3-AS1 could be negatively regulated by SNAI2, loss-of-function and gain-of-function studies regarding SNAI2 were further performed in two GC cell lines. The qPCR analysis showed that ELF3-AS1 and ELF3 were significantly downregulated in the SNAI2 overexpression cell lines but were significantly upregulated in the SNAI2-depletion cell lines (Fig. 1C-E). These results suggested that both ELF3-AS1 and ELF3 were negatively regulated by SNAI2 in GC.
ELF3-AS1 is an antisense lncRNA (head-to-head) of the epithelial tumor suppressor gene ELF3. Promoter analysis revealed that ELF3-AS1 promoter contained the sequence “GGTACAGGTGGGT”, which was predicted to be recognized by both SNAI2 and SNAI1. This sequence was located 829bp upstream of the ELF3-AS1 transcription start point, which was also the junction point between exon 2 and intron 2 of the ELF3 gene (Fig. 1F). Thus, we speculated that both ELF3-AS1 and ELF3 might be transcriptionally regulated by SNAI2 and SNAI1.
ELF3-AS1 and ELF3 were transcriptionally repressed by both SNAI2 and SNAI1.
LncRNA ELF3-AS1 is abundant in human cells and can be effectively captured by magnetic beads when RNA sequencing. In order to make our results more convincing, we used RNA-seq studies to visualize the expression of ELF3-AS1 and ELF3. As expected, RNA-seq data showed that ELF3-AS1 and ELF3 transcripts were both greatly reduced in SNAI2 (or SNAI1) overexpression cell lines (Fig. 2A-F). Besides, the inhibitory intensity of SNAI2 on the expression of ELF3 and ELF3-AS1 was much greater than that of SNAI1 (Fig. 2B, E).
Furthermore, to figure out whether the negative regulation of ELF3 and ELF3-AS1 by SNAI2/SNAI1 occurred at the transcriptional level, dual-luciferase assays and CHIP assays were performed in the SNAI1 and SNAI2 overexpression cell lines. As shown in Fig. 2G, when the SNAI1/2 binding sequence was mutated, the strong repression of SNAI1/2 overexpression on luciferase expression could be partially restored, suggested that this sequence was necessary for SNAI1/SNAI2 to inhibit ELF3/ELF3-AS1 expression. On the other hand, CHIP assays showed that SNAI1 and SNAI2 could directly bind to the ELF3-AS1 promoter (Fig. 2H-J). Taken together, both SNAI2 and SNAI1 could transcriptionally repress the expression of ELF3 and ELF3-AS1 in GC.
The reduced ELF3-AS1 and ELF3 expression were clinically associated with poor prognosis in GC.
The ELF3-AS1 expression pattern in 30 pairs of GC tissues was determined by qPCR assay. As expected, ELF3-AS1 was down-regulated in more than 80% of GC samples compared to the corresponding normal samples (Figure. 3A). On the other hand, we analyzed the correlation between level of ELF3-AS1 expression and the clinical characteristics of GC tissues from the cancer genome atlas (TCGA, n = 375) database. LncRNA ELF3-AS1 was low expressed in diffuse and poorly differentiated gastric cancer tissues (Fig. 3B and C, p < 0.0001). However, no significant changes in ELF3-AS1 expression levels were observed between GC patients with different TNM stages (Fig. 3D-F, p > 0.05). Overall survival analysis showed that GC patients with lower expression level of ELF3-AS1 possessed shorter overall survival time (Fig. 3G, p = 0.029).
Our previously published study had implied that transcription factor ELF3 plays tumor-suppressive roles in GC.(11, 13) Herein, we further confirmed that ELF3 protein and ELF3 mRNA were significantly downregulated in GC (Fig. 3H-J). Besides, lower expression of ELF3 was observed in the diffuse GC compared with intestinal GC (Fig. 3K). The low expression of ELF3 was positively correlated with the malignant progression of GC (Fig. 3L and M). Moreover, patients with lower ELF3 expression had a poorer overall survival time and disease-free survival time in the GSE62254 GC cohort (Fig. 3N and O).
According to the clinical outcomes, both ELF3-AS1 and ELF3 functioned as tumor suppressors in GC. Since ELF3-AS1 and ELF3 were transcriptionally repressed by SNAI2 and SNAI1, we further analyze the expression signatures and prognostic values of SNAI2 and SNAI1 in GC. As expected, SNAI2 and SNAI1 were upregulated in GC (Figure S1A and B). Besides, the overexpression of SNAI2 and SNAI1 predicted poor prognosis in the two independent GC cohorts (Figure S1C-F).
ELF3 negatively regulated GC cell metastasis but cannot regulate ELF3-AS1 expression
Protein coding genes are usually highly co-expressed with their neighboring lncRNAs(14). Similarly, ELF3-AS1 and ELF3 were confirmed to be highly co-expressed in the normal stomach tissues, GC cell lines and tissues (Fig. 4A-D, p < 0.0001). Interestingly, when the ELF3-AS1 was effectively knocked down in GC cell lines, the transcripts of ELF3 were significantly reduced to approximately 60–70% (Figure. 4E-H). However, it’s not clear how ELF3-AS1 affects ELF3 expression in GC.
Given ELF3 belongs to ETS transcription factor family, we initially assumed that the co-expression of ELF3-AS1 and ELF3 may be due to ELF3 regulating the expression of ELF3-AS1. To verify this possibility, loss-of-function and gain-of-function studies regarding on ELF3 were performed in two GC cell lines. However, the expression of ELF3-AS1 had no significant alteration after knockdown or overexpressing of ELF3 in GC (Figure. 4I and J). Although transcription factor ELF3 cannot regulate ELF3-AS1 expression, the scratch wound healing assays and transwell assay still confirmed that ELF3 negatively regulated GC cell migration and invasion (Figure. 4K and L).
ELF3-AS1 mainly inhibited GC metastasis through repressing SNAI2 signaling
To determine whether ELF3-AS1 has a tumor-suppressive effect in GC, loss- and gain-of-function studies were performed in GC. Cell apoptosis assay showed that depletion of ELF3-AS1 remarkably accelerated early apoptosis of GC cells (Figure S2A). Besides, cell proliferation, transwell and scratch wound healing assays showed that ELF3-AS1 knockdown promotes GC cell proliferation, migration and invasion (Figure. S2B-F). The overexpression of ELF3-AS1 significantly inhibited the proliferation, migration and invasion ability of HGC-27 cells (Figure. S2G-I). Moreover, we examined the effect of ELF3-AS1 silencing in a xenograft GC model in vivo. The tumor growth of GC cells silencing ELF3-AS1 was significantly increased when compared to that of the control GC cells (Figure. S2J and K). These results together suggested that ELF3-AS1 inhibited proliferation and metastasis of GC cells in vitro and in vivo.
We analyzed the differentially expressed miRNAs after ELF3-AS1 knockdown by miRNA sequencing (Fig. 5A). Surprisingly, among the top 10 miRNAs that were most significantly down-regulated after ELF3-AS1 knockdown, miR-33a, miR-33b and miR-203a were reported to be able to target SNAI2 expression (Fig. 5B).(15–17) To confirm the reliability of miRNA sequencing, we examined the expression level of miR-33a, miR-33b and miR-203a in the ELF3-AS1 knockdown cell lines by qRT-PCR assays. The results showed that knockdown of ELF3-AS1 can indeed significantly down-regulate the expression of miR-33a, miR-33b and miR-203a (Fig. 5C-E). In turn, the depletion of ELF3-AS1 led to significant up-regulation of SNAI2 mRNA and protein (Fig. 5F-I).
In addition, the gene expression profiles of SNAI2 overexpression and ELF3-AS1 knockdown were very similar (Figure S3A). The genes strongly regulated by SNAI2 overexpression also have similar expression changes in ELF3-AS1-depleted GC cells (Figure S3B-D). For examples, the genes strongly inhibited by SNAI2 overexpression, such as IGFBP3, ITGB4, TJP3, PPL and DDIT4, were also significantly down-regulated in the ELF3-AS1-depleted GC cell lines. The genes strongly induced by SNAI2 overexpression, such as IL11, THBS1, INSL4 and linc02104, were also remarkably upregulated in the ELF3-AS1-depleted GC cell lines. These results strongly implied that knockdown of ELF3-AS1 could not only upregulate SNAI2 expression but also greatly activate the downstream signaling of SNAI2 in GC.
Based on above findings, we speculated ELF3-AS1 may inhibit GC progression through repressing SNAI2 expression. To verify this possibility, the rescue assays were performed in two different GC cell lines that stably knocked down ELF3-AS1. The results showed that exogenous siRNA targeting SNAI2 rescued the tumorigenic properties of ELF3-AS1-depleted GC cell lines (Fig. 5J and K). These data strongly indicated that ELF3-AS1 mainly inhibited the migration and invasion of GC cells through repressing SNAI2 signaling.
The nuclear-localized lncRNA ELF3-AS1 plays critical roles in the cell cycle progression.
The biological function of lncRNA is closely related to its subcellular location.(18) ELF3-AS1 was a nuclear-localized lncRNA in GC (Fig. 6A and B). Previous study had reported that ELF3-AS1 was a cell cycle-related lncRNA.(19) Our study also showed that lncRNA ELF3-AS1 played essential roles in cell cycle progression. Silencing ELF3-AS1 significantly accelerated the G1/S transition of the cell cycle in GC (Fig. 6C and D). Interestingly, RNA-seq analysis showed that knockdown of ELF3-AS1 resulted in a significant up-regulation of almost all histone-coding genes by more than 2 times (Figure. 6E and F). It’s well known that the synthesis of histones is mainly in the S phase of cell cycle, which is synchronized with DNA replication. These results indicated that ELF3-AS1 negatively regulates cell cycle progression of GC cells by affecting G1/S transition and histone synthesis.
To better understand of the molecular mechanism of ELF3-AS1 in regulating the cell cycle process, we analyzed the expression changes of cell cycle-related genes after knockdown of ELF3-AS1. The results showed that knockdown of ELF3-AS1 increased the expression of CDK6 and CASP7, while decreased the expression of CDKN1A (also known as p21) in GC (Figure. 6G and H). The p21 proteins functioned as a cell cycle checkpoint of G1/S transition.(20) The CDK6/CCND1 protein complex is very important for cell cycle G1 phase progression and G1/S transition.(21) Therefore, we considered that the promotion of G1/S transition caused by ELF3-AS1 knockdown may be due to the downregulation of P21 and the upregulation of CDK6.
ILF2/ILF3 complex could directly modulate the stability of ELF3-AS1/ELF3 RNA duplex.
The potential proteins interacted with lncRNA ELF3-AS1 were identified by RNA pull-down analysis (Fig. 7A). According to the mass spectrometry (MS) analysis of the differential bands located at 45 Daltons (Fig. 7B), there were 7 proteins with matching coverage greater than 20% (Table S4). 3 of them, RINI, ILF2 (also known as NF45) and TARBP2, were double-strand RNA binding proteins (Fig. 7C). Interestingly, another protein named ILF3 (also known as NF90/NF110) was also appear in the MS results of this band (Fig. 7D and E). Our subsequent western blot assay further verified that ILF2, ILF3 and TARBP2 could bind to exogenous lncRNA ELF3-AS1 (Fig. 7F). Moreover, CHIRP pulldown assay verified that endogenous ELF3-AS1 also bound to the ILF2 and ILF3 proteins (Fig. 7G and H). The Venn plot showed that approximately 22 proteins are displayed at the intersection of the three MS results, including ILF2, ILF3, RINI, etc. (Table S4-6, Fig. 7G and H). RNA immunoprecipitation assay showed that the ELF3-AS1 transcripts bound to ILF2 and ILF3 proteins were thousands of times higher than the control IgG group (Fig. 7I-K). To figure out which segment of ELF3-AS1 transcript could interact with ILF2 and ILF3, we truncated ELF3-AS1 transcripts of different lengths (Fig. 7L). The results showed that the first exon of ELF3-AS1 was necessary for the interaction between the ELF3-AS1 and the ILF2/ILF3 complex (Fig. 7M and N).
The ELF3 gene has many different transcripts due to alternative splicing. Among those different types of ELF3 transcripts, ELF3-201, ELF3-202 and ELF3-203 can encode full-length ELF3 protein. It’s worth mentioning that the ELF3-201 transcript and the ELF3-AS1 transcript overlapped by about 664 nucleotides (Fig. 8A). In other words, ELF3-AS1 can combine with the first exon region of ELF3-201 to form a double-stranded RNA molecule. On the other hand, RNA-seq analysis revealed that knockdown of ELF3-AS1 had a more profound effects on ELF3-201 expression compared to the ELF3-203 or any other ELF3 transcripts level (Fig. 8B). These data implied that the ILF2/ILF3 complex may bind to the double-stranded RNA formed by ELF3-AS1 and ELF3-201. To further verify this probability, we also examined the ELF3-201 transcript level in the RIP assays of ILF2/ILF3. The ELF3-201 transcripts bound to ILF2 and ILF3 were much higher than the control IgG group (Fig. 8C-E), suggesting ILF2/ILF3 complex could bind to the double-stranded RNA formed by the first exons of ELF3-AS1 and ELF3-201.
It has been reported that ILF2/ILF3 complex play roles in regulating mRNA stability.(22) The RNA-seq data and the qPCR assays indicated that knockdown of ILF3 significantly decreased the mRNA level of ELF3-AS1 and ELF3-201, while knockdown of ILF2 significantly increased the mRNA level of ELF3-AS1 and ELF3-201 (Fig. 8F and G, Figure S4A-I). These results suggested that ILF2 and ILF3 protein possessed opposite effects on the stability of ELF3-AS1 transcripts. Previous study reported that NF45 functions as a regulatory subunit in ILF2/ILF3 complexes.(23) Interestingly, we also noted that knockdown of ILF2 could obviously affect the alternative splicing of ILF3 gene (Fig. 8H). qRT-PCR assays showed that knockdown of ILF2 significantly upregulate the expression of NF90, but significantly decreased the expression of NF110 (FigureS4J and K). Therefore, we speculated ILF2 (NF45) might regulate ELF3-AS1 and ELF3-201 expression through affecting the alternative splicing of ILF3 gene.
{kind=link}