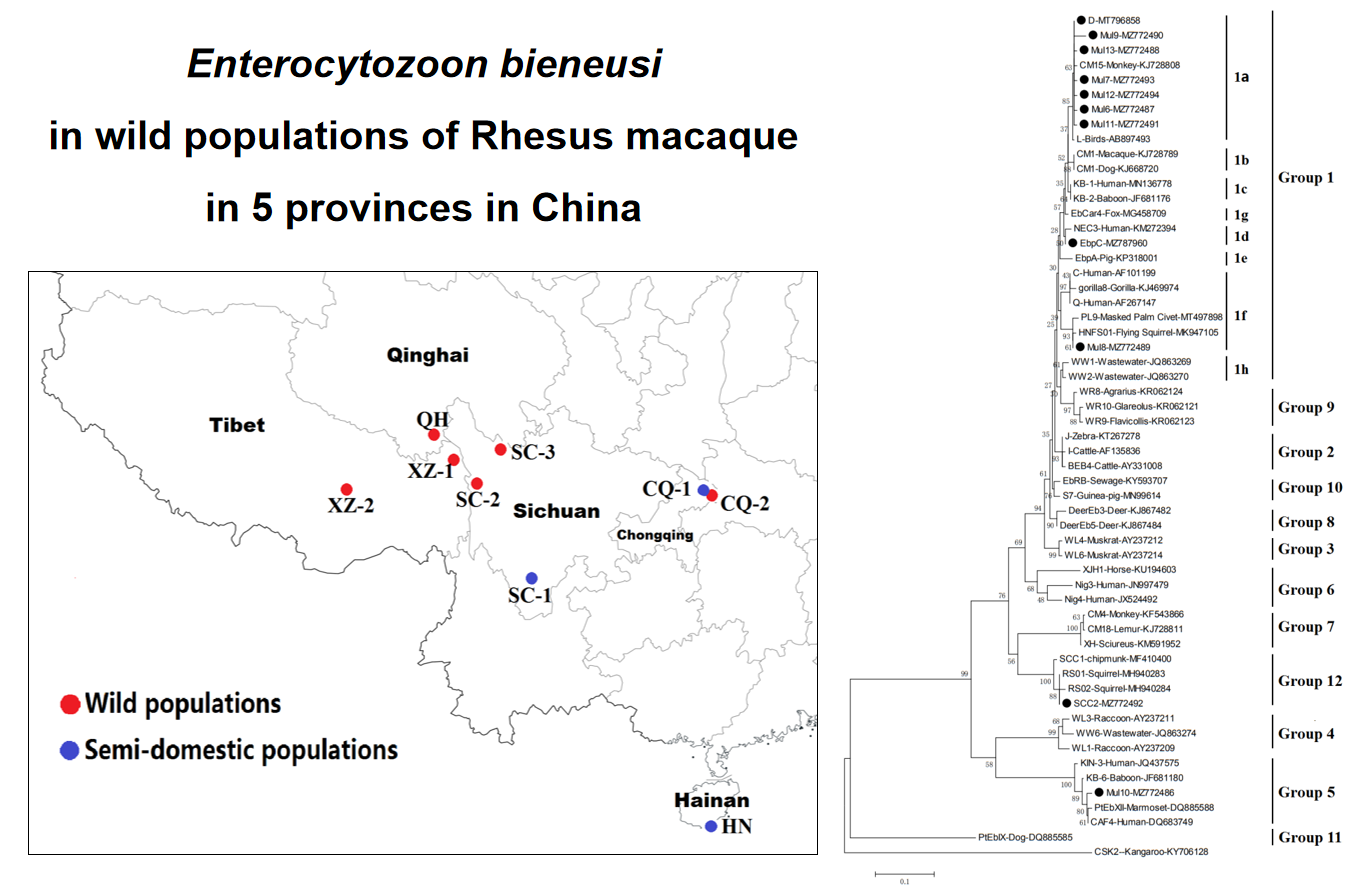
E. bieneusi is the most diagnosed species in the 17 species of microsporidia which reported to cause infections in humans, domestic and wild animals[3]. The first documented transmission of E. bieneusi infection in human (afflicted with AIDS) and a Rhesus macaque (afflicted with simian immunodeficiency virus) was reported in 1997 [16]. At present, the research of E. bieneusi in wild or captive non-human primates (NHPs) has been reported all over the world. In China, researches in Rhesus macaque found that the prevalence range of E. bieneusi is 4.8 to 56.5%, these studies mainly focus on captive populations[13, 17, 18]. In our study, we find out the infection of E. bieneusi in 6 wild and 3 semi-domestic populations of Rhesus macaque from 5 provinces in China (i.e. Sichuan, Tibet, Qinghai, Chongqing and Hainan). These populations, SC-2, XZ-1, XZ-2, HN and CQ-1 had high infection rates 10.3–26.3%. Whereas, CQ-2 (6.7%) and SC-1(6.9%) had low infection rates. Food-borne transmission of E. bieneusi has been documented, although the contamination of vegetables and fruits with this pathogen was also reported in China[19]. The location of the four populations SC-2, SC-3, XZ-1 and XZ-2 were close to villages and pastoral areas, where Rhesus macaque had the opportunity to consume the same water and food that the villager and free-rang livestock were used. We consider that it may cause parasitic disease among humans, livestock and wild Rhesus macaque. Previous study reported that E. bieneusi has spread infection in pigs, yaks and cattle in Tibet China [20–23], it is correlated to our study. Another study in eastern Qinghai found E. bieneusi in free-rang sheep and yaks[24], but in our study, we didn’t find the infection in QH (n = 40) southern Qinghai population. This population was living in primitive forests and had little or no contact with villagers and livestock. While, CQ-2 population (6.70%) had the lowest infection rate was observed to very close with the cliffs on both sides of the Yangtze River, there may be no contact with other potential infection sources of E. bieneusi except water source. In the semi-wild populations, infection rate of HN population (15.2%) was almost same with the previously study in the Nanwan Monkey Island (15%)[14]. The 3 semi-wild population (CQ-1. SC-1 and HN) were living in the natural scenic spot with many tourists. The populations CQ-1 and SC-1 had similar geographical and climatic conditions, which were different from HN. But the infection rate of SC-1 (6.9%) was lowest than HN (15.2%) and CQ-1 (15.9%). In previous research the infection of E. bieneusi in 11 captive Rhesus macaque populations showed the significant result of infection rate in different zoos. It could be mainly due to the different environment and level of management in the zoos[17]. In the semi-wild population living in the natural scenic spot, the difference of infection rate may also be affected by many factors (i.e. Health status of the host, detection methods, sample size and the experimental design).
In our study, genotype D was the most prevalent and observed in 7 populations with the highest infection rate (63.15%). While, another known genotype EbpC was only found in XZ-2 population. Genotype D and EbpC were commonly found in the humans, domestic and wild animals all over the world [2]. In China, genotype D (synonyms: PigEBITS9, WL8, Peru9, CEbC, and PTEb VI) has been detected in infants, HIV positive patients and HIV negative patients[3, 25–28]. Genotype EbpC (synonyms: CHG23, E, Peru4, SC03, WL13 and WL17) was commonly found in humans and more than 15 animal species[15, 29–32]. According to our phylogenetic analysis, the novel genotypes, Mul6, Mul7, Mul9, Mul11, Mul12 and Mul13, were genetically closely related to the genotype D, and all were clustered in Group 1-a. Whereas, the novel genotype Mul8 was distributed in Group 1-f. At present study, most genotypes in Group 1 are host adaptation and zoonotic[2]. Therefore, we conjecture these new genotypes in Group 1 may be zoonotic. Genotype SCC-2 (n = 1) was found in population SC-1. Genotypes SCC-1 ~ 3 and RS01 were clustered in Group12[33], these genotypes were found only in rodents from China in previous studies[34, 35]. Our study is the first record to find SCC-2 in NHPs. It may transmit from infected rodents to Rhesus macaque. Further studies are needed to understand the host range and public health importance of genotype SCC-2. Moreover, the novel genotype Mul10 may be host-specific, it was clustered in Group 5 with the host-specific genotypes CAF4, KB-6, KIN-3 and PtEb-XII which have been found only in those hosts from which they were originally reported[36–39].
It is noteworthy that the 4 wild populations (SC-2, SC-3, XZ-1 and XZ-2) which found genotypes D, EbpC, and the potentially zoonotic novel genotypes (Mul7, Mul8, Mul9, Mul11 and Mul12) were located close to villages and pastoral areas, it induces a great risk of E. bieneusi infection in humans and free-range livestock. Besides that, the Rhesus macaque from 3 semi-wild populations, had genotype D and potentially zoonotic novel genotypes (Mul3 and Mul6), contained more opportunities to contact with tourists and managers of scenic spot. Considering the density of the crowd, there is always a risk of parasite transmission between humans and monkeys,
{kind=link}