Regulation of wheat grain weight potential by the maternal grain tissues during development has been previously suggested and investigated by other groups [10, 13, 18], but to our knowledge, this work represents the first example of this interaction being explored under high p.a. temperatures. Using RNA-Seq analysis and cross-referencing with existing data sets [23, Pellny published online: Arrayexpress - E-MTAB-8397] we were able to better characterise the effect of high p.a. temperatures on the expression profiles of genes predominantly expressed in either the pericarp or endosperm and relate these genes to particular cellular mechanisms. We have focused in particular on genes involved in cell wall restructuring, since our driving hypothesis was that high temperatures, when experienced p.a., would disproportionally accelerate the rate of development of the pericarp tissues compared to the endosperm, resulting in a premature decrease in the ability of pericarp cell walls to expand. This, in turn, would limit endosperm expansion, and therefore grain size.
High p.a. temperature exposure reduces mature grain weight in cv. Cadenza
The early grain filling phase (6daa-10daa) has been the focus of previous studies investigating the effect of high p.a. temperature on wheat, with high p.a. temperatures during this developmental stage having been reported to result in reductions in mature grain weight [25, 26, 27, 28, 29]. The results of Experiment 1, in this study, show that high p.a. temperatures applied towards the end of the celluarisation phase of grain development (around 6daa) for 4-days or longer reduce mature grain weight in wheat, whilst a 2-day high p.a. temperature treatment was insufficient to produce significant differences. This is in contrast with the results of Talukder et al. [29], who observed a reduction in mature grain weights in 4 commercial Australian bread varieties subjected to a single day’s exposure of high p.a. temperature treatment (35 °C) between 7 and 10daa, and highlights the genotypic variability in the response of wheat cultivars to high p.a. temperatures.
The results of Experiment 1 also suggest that there is not a proportional effect between the duration of a high p.a. temperature treatment and the reduction in mature grain weight: the reduction in mature grain weight observed in plants that had undergone 4-days of treatment was not significantly different to those observed in plants that experienced 6, 8, 10 and 12 days of high p.a. temperature treatment. This result supports suggestions by Stone et al. [30] that when brief periods of high p.a. temperature occur, yield potential in wheat is decreased by a more or less a fixed extent. The results of this experiment also support the findings of Nuttall et al. [31] who observed that the application of 4-days of cumulative exposure to a high p.a. temperature treatment of 35/15 °C from 2daa for 6 hours per day resulted in 15% reduction in individual grain weight in cv. Yipti and that this reduction plateaued from this point onward with further durations of treatment. Therefore, 4-days of high p.a. temperature treatment, either applied consecutively or days apart, would represent a threshold beyond which sensitivity to high p.a. temperature declines, and possibly suggests that the high temperature treatment may trigger the imposition of a structural limit on the sink capacity of the grain.
Although maternal tissue control of seed size and endosperm sink capacity has been well studied [17, 32], the mechanism via. which this is achieved has not so far been established. High p.a. temperature treatment, administrated as reported in the experiments here discussed, induced significant down-regulation of pericarp genes with a known role in regulating or modifying cell wall expansion. The down regulation of these genes at around 10daa following the same treatment duration that was demonstrated to result in a reduction in mature grain weight in Experiment 1 adds further support to the hypothesis that high p.a. temperature induces modifications to the expansion capability of the pericarp, which may constrain endosperm cell expansion and present a degree of control over final grain size and weight.
High p.a. temperatures reduce the ability of the grain to accumulate water
Water accumulation is the main driver for endosperm cell expansion and grain moisture content is therefore closely associated with the developmental stage of the wheat grain [20, 33]. Furthermore, maximum moisture content of the wheat grain has been reported as a reliable indicator of maximum grain volume [34] and, therefore, of final grain weight [35]. Our study showed that high p.a. temperatures affects the potential for water accumulation in the grain: maximum grain moisture content was significantly lower in high p.a. temperature treated grains compared to control grains and this maximum was reached 4-days earlier at 14daa. Pericarp degeneration, detectable as a loss of greenness, has been previously been shown to occur prior to the termination of rapid net water accumulation in wheat grain [36]. In addition, the degeneration of the pericarp was postulated to be causally related to the achievement of the max wheat grain sink capacity, on the basis that the end of endosperm cell division and the cessation of net water accumulation closely coincide [37].
At the transcriptomic level, in this same time interval (6–14 daa), high p.a. temperatures induced down-regulation of genes predominantly expressed in the pericarp and involved in cell wall expansion in grains sampled 10daa, prior to the stabilisation of maximum water content. In support of this observation, Lizana et al. [10] also reported a down-regulation in the expression of expansin transcripts just prior to the stabilisation of grain moisture content. Our result suggests that a premature loss of cell wall plasticity in the pericarp under high p.a. temperature may be at the basis of the reduced volumetric capacity observed in wheat grain exposed to high p.a. temperature.
Pericarp gene expression under high p.a. temperature
Studies by Altenbach and Kothari [38] and Wan et al. [20], showed that high p.a. temperature accelerate the normal pattern of gene expression within the wheat grain. In our study, where treatment was applied from 6daa, we observed the largest number of DEGs between high p.a. temperature treated and control samples at 10daa, after only 4-days of treatment, rather than at the later time point of 14daa. In agreement with the findings of the above cited authors, PCA showed that high p.a. temperature accelerates gene transcription patterns associated with development, exemplified by lower PC1 values (lower PC1 value = Increased developmental stage) for high p.a. temperature treated samples than control samples. Furthermore, PC2 separated control and high p.a. temperature treated samples along the Y-axis, suggesting that temperature can affect gene expression by more than simply accelerating development e.g. heat stress responses.
Among the DEGs, we identified 12 predominantly pericarp expressed genes showing both significant differences in expression between 10 and 14daa and a significant interaction with heat treatment. These included 5 LTP genes significantly down-regulated at 10daa in samples exposed to high p.a. temperature, with three of these being subsequently up-regulated at 14daa and two remaining at similar expression levels between the two sampled stages. LTPs are a group of highly divergent, small basic proteins involved in cutin development, cell defence, the water permeability of cell membranes and cell wall extension [39, 40, 41].
Of the 5 genes encoding LTPs in our data, 3 of those genes encoded non-specific LTPs (nsLTPs), and members of the nsLTP family have been showed to be responsive to one or multiple abiotic stressors (reviewed in Liu et al. [42]), including drought [43, 44, 45, 46], salt [44, 45], low temperature [45, 47], and high temperature [45]. Transcript levels for nsLTPs in wheat have been previously shown to increase from around 12daa until the late stages of grain development [48] under high p.a. temperatures (37/28 °C). Also in our study, nsLTPs encoding genes were significantly differentially expressed between developmental stages and showed a significant interaction with temperature. We observed down-regulation of three NSLTPs at 10daa under high p.a. temperatures compared to expression of these genes at 6daa and 10daa under control temperatures, followed by up-regulation at 14daa. We suggest this pattern of expression may reflect nsLTPs multiple functions: these genes may be down-regulated under high p.a. temperature conditions at 10daa reflecting a general reduction in pericarp cell wall extension consistent with the role of nsLTPS as cell wall plasticisers [40] before being upregulated at 14daa in response to high temperatures, possibly to facilitate cuticle development as part of an earlier maturation response of the grain.
Peroxidase
Another gene differentially expressed between 10 and 14daa with a significant interaction with temperature treatment was PER2/PER42, a gene encoding an un-specified haem-b class III Peroxidase and which was down-regulated in high p.a. temperature treated samples at 14daa. Class III peroxidases are secreted glycoproteins known to be involved in a broad range of physiological processes, including regulation of cell elongation and cell wall modification [49, 50]. They are able to promote rigidity of cell walls by forming strong intracellular bonds by oxidation of aromatic cell wall compounds (e.g. lignin), but they can also generate radical oxygen species (ROS) that can break covalent bonds in cell wall polymers such as hydroxyl radical (OH°) [51, 52]. As such, they perform dual functions within plant cell walls, regulating both cell wall stiffening and relaxation. Lipid peroxidation under high temperatures has been reported during the grain-filling stage in wheat, with high temperatures being associated with increase in membrane damage and decrease antioxidant levels [53]. Also in wheat there is evidence of down-regulation of the activity of antioxidant enzymes like peroxidases occurring under high temperatures [54] and exacerbating the accumulation of ROS. The downregulation of the PER2/PER42 gene observed in our study could be linked to ROS accumulation under heat stress, either as a cause or a response.
Gene ontology and functional analysis
Knetminer analysis
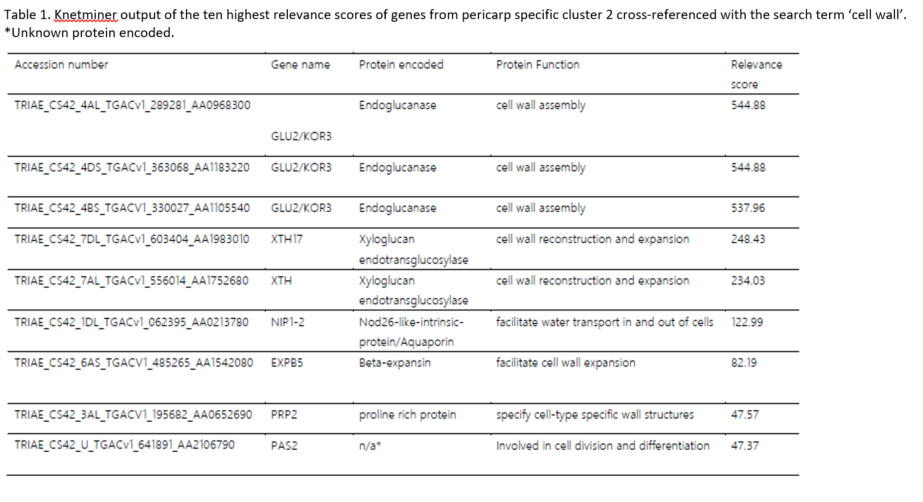
Knetminer webtool analysis with the search term ‘cell wall’ applied to the down-regulated genes within pericarp cluster 2 identified endoglucansases, xyloglucanase, an aquaporin and β-EXP. Endoglucanases are a group of membrane-bound enzymes that hydrolyse cell wall polysaccharides with contiguous (1, 4)-β -glycosyl residues in their chain and are implicated in the breakdown of plant cell walls during development [55]. The KORRIGAN (KOR) gene, a specialised member of the endoglucanases, has been shown to be expressed in the expanding cell walls of A.thaliana [56], with one member of the KOR family, the KOR3 gene, in particular, having been associated with cell wall biosynthesis [57]. In our study, we report the down regulation of three KOR3 genes in the pericarp under high p.a. temperature, which suggests a reduction in cell wall assembly in the outer layers of the grain and, consequently, in their expansion potential.
Within the pericarp cluster 2 genes, the Knetminer analysis webtool also identified members of the XTH family. Some XTHs have been shown to be down-regulated under drought conditions in Barley’s awns [58], however no study has reported so far on the expression of XTH genes in wheat grain under high p.a. temperature. Our data shows that whilst XTHs were down regulated within the pericarp in both control and treated samples between 10 and 14daa, the reduction in the expression of these genes was greater under high p.a. temperatures, which also suggests a reduction in the expansion capacity of the cell walls within the pericarp. Finally, EXPB5 was also identified from the pericarp cluster 2 genes by the Knetminer webtool when searching with the term ‘cell wall’. EXPB5 encodes a β-EXP, belonging to a class of cell wall proteins which have previously been shown to be crucial in regulating cell wall restructuring via the disruption of hydrogen bonds between cellulose microfibrils and cross-linking glycans, thus allowing cellular expansion under turgor driven pressure [59, 60].
Expression of α- EXP genes derived from wheat grain pericarp RNA has been reported to peak between 8-12daa [61] and to rapidly decline after 25daa [10]. Whilst these studies show the importance of α-EXP genes in the early stages of wheat grain development, there is less information regarding the function and presence of β-EXPs in wheat grains in more advanced stages of development.
α and β EXPs make up the two largest families of EXP genes and while they share only around 20% amino-acid identity [60], members from both families appear to destabilise noncovalent load bearing bonds between cellulose microfibrils in plant cell walls [62].
OsEXPB5, a member of the β-EXP family in rice, has been showed to be involved in root hair growth [63]. While there is little information pertaining to the specific action of EXPB5 in wheat grain cell walls, its presence in pericarp cluster 2 suggest that the expression pattern of this cluster may be representative of changes in the cell wall of the pericarp under high p.a. temperatures: down-regulation of EXPB5 between 10 and 14daa is stronger in high p.a. temperature treated samples, suggesting a higher increase in the rigidity of the pericarp cell walls of these grains compared to untreated samples.
GO term analysis
GO term enrichment for pericarp cluster 2 revealed that the most significant GO term was ‘cuticle development’ (GO id: 0042335). The cuticle acts primarily as a permeability barrier involved in regulating water loss and protecting plant organs from dirt, bacteria and other microorganisms [64]. During the early stages of anthesis the developing wheat grain contains a number of osmophilic cuticles or cuticular membranes but most of them are gradually removed p.a. by enzymatic degradation [65] so that only two cuticles persist in the developing grain up to maturity: one being found on the outermost layer of the grain, the outer-epidermis, and the other being positioned on the surface of the nucellar epidermis.
The downregulation of genes contained within pericarp cluster 2 at 10daa in high p.a. temperature treated samples compared to control samples indicates a clear effect of high p.a. temperature treatment on the expression of genes involved in cuticle development. Previous studies have shown that the cuticle of wheat leaves and its specific wax composition is important in regulating drought tolerance in different cultivars [66]. The outer-epidermis cuticle of the wheat grain forms from around 7daa and reaches is maximum thickness by 17daa [65], after which it remains structurally unchanged until maturation apart from becoming appressed to the inner cuticle. Genes involved in the formation of the cuticle are therefore expected to be down-regulated from around 14-17daa as the cuticle ends its development. We observed down-regulation of genes contained within pericarp cluster 2 and annotated as involved in ‘cuticle development’ from 10daa in high p.a. temperature treated samples, which supports the hypothesis of an acceleration in the development of the pericarp under high p.a. temperature. Given the cuticle’s important role in regulating moisture loss in plant tissues, increased expression of genes involved in cuticle development might be expected in order to reduce water losses at high temperature, but in fact the opposite was observed with a more rapid decrease of these transcripts. Possibly this reflects importance of cuticular transpiration in cooling tissues to maintain function.
The most significant GO term derived from the genes contained within endosperm predominant cluster 3 was ‘The killing of cells of other organisms’ (GO id: 0031640). The genes contained within endosperm predominant cluster 3 experienced increased upregulation under high p.a. temperatures compared to the control samples. Previous studies have shown a large degree of functional overlap in defence-associated genes that respond to both biotic and abiotic defence mechanisms [67]. In addition, a transcriptome microarray analysis of grains from a Chinese bread wheat cultivar (Jimai 20), reported in Yu et al. [68] showed that genes associated with defence against both biotic and abiotic stresses were all up-regulated during the grain filling period, particularly between 11-15daa; the results obtained from our experiment, whereby defence associated genes are up-regulated to a high degree between 10 and 14daa in high p.a. temperature treated grain are in agreement with the above mentioned work. Therefore, it appears that high p.a. temperatures accelerate the expression profile of genes involved in cuticle development in the pericarp (down-regulated) and defence genes within the endosperm (up-regulated) between 10 and 14daa.
{kind=link}