This study has been reported in accordance with the ARRIVE guidelines for transparent preclinical reporting. Baseline characteristics of the dose validation study were compared between control and melphalan groups with no significant differences identified (Table S1).
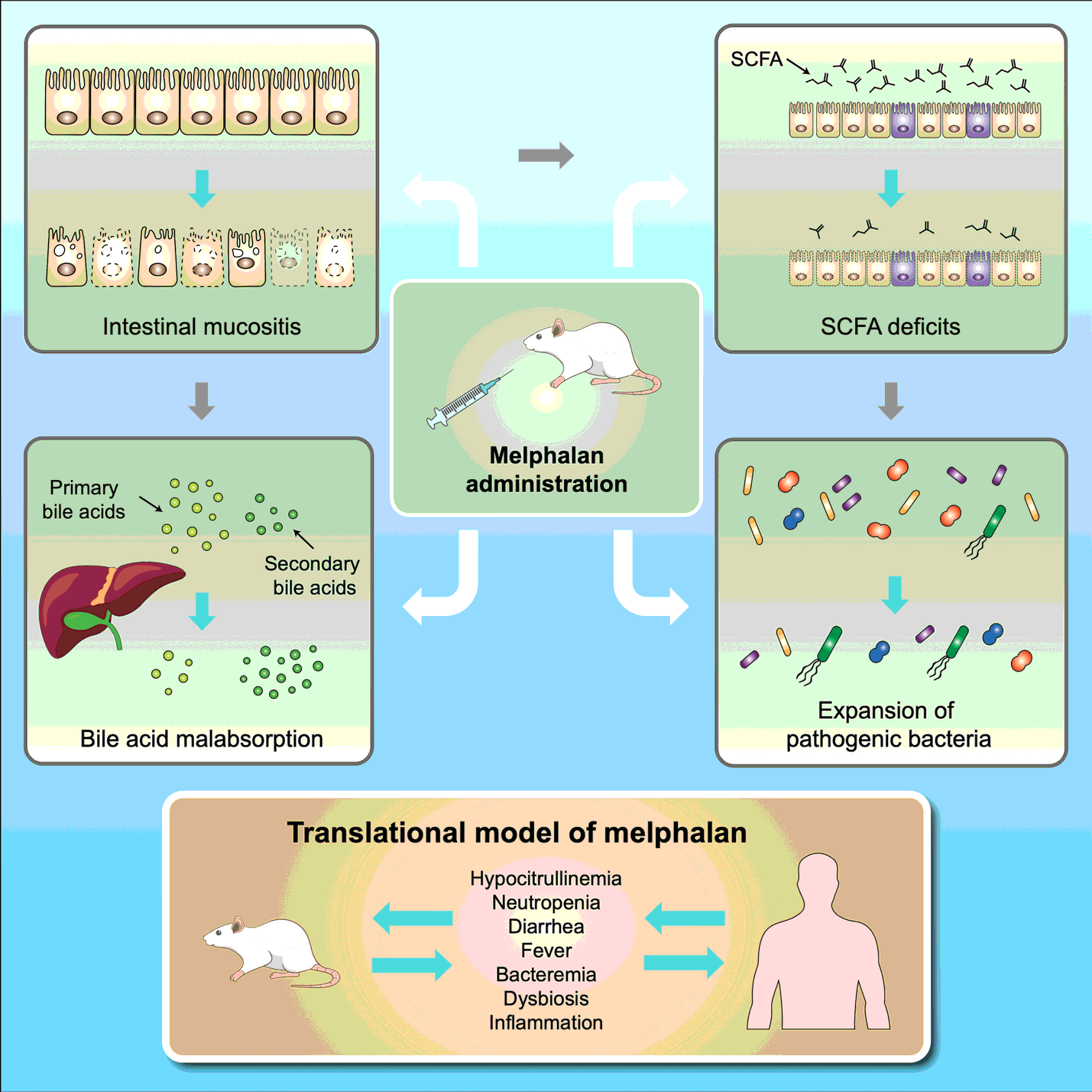
Intravenous melphalan induces a dose-dependent toxicity phenotype characterized by weight loss, diarrhea and mucosal barrier injury
Melphalan caused severe toxicity at doses > 6 mg/kg with 100% mortality observed in rats treated with the highest dose (8 mg/kg; Fig. 1A). Melphalan caused dose-dependent weight loss (Fig. 1B), with maximum weight loss observed at day 4 at self-limiting doses (Fig. 1C). Melphalan caused a rapid decrease in plasma citrulline, which was lowest at day 4 (15.25 ± 1.06µM). Citrulline was strongly correlated with weight loss at day 4 (R2 = 0.9927, P < 0.0001), validating its applicability as a biomarker of mucosal barrier injury (Fig. 1D-G. Hypocitrullinemia (< 10µM) was evident at doses of 6 mg/kg and 8 mg/kg, however these doses were also accompanied by unacceptable diarrhea severity and mortality (100% for 8 mg/kg, Fig. 1H-L). Melphalan administered at 5 mg/kg caused self-limiting mucosal barrier injury, with transient grade 1 diarrhea peaking at day 7 (Fig. 1J).
5 mg/kg intravenous melphalan causes a biphasic, self-limiting clinical phenotype accompanied by fever and severe neutropenia
Melphalan at 5 mg/kg caused significant weight loss compared to vehicle control animals (-6.4 ± 1.01% relative to baseline, P < 0.0001 Fig. 2A/B). Significant decreases in citrulline were evident on day 2 (84.1 ± 1.9µM vs 37.6 ± 1.3µM, P < 0.0001), day 4 (81.5 ± 2.4 µM vs 12.8 ± 1.24µM, P < 0.0001) and day 7 (78.6 ± 2.7µM vs 59.2 ± 14.0µM, P = 0.012), returning to baseline by day 10 (Fig. 2C). In melphalan treated animals, weight loss and citrulline were moderately correlated (R2 = 0.433, P < 0.0001). Melphalan also caused a significant reduction in food consumption, which was significant from day 1–7 (Fig. 2E), however this was not accompanied by any change in water intake (Fig. 2F).
Body temperature spiked at day 6 in melphalan-treated animals (-0.14 ± 0.10oC vs + 0.70 ± 0.09oC, P < 0.0001, Fig. 2G). Melphalan also induced severe leucopenia, with a significant decrease in total WBC count at day 4 (6.1 ± 0.8 × 109/L vs 1.0 ± 0.2 × 109/L, P = 0.03, Fig. 2H), day 7 (4.8 ± 0.1 × 109/L vs 0.7 ± 0.1 × 109/L, P < 0.0001, Fig. 2H) and day 10 (5.2 ± 0.4 × 109/L vs 3.0 ± 0.6 × 109/L, P = 0.02, Fig. 2H). This was explained by significant ablation of neutrophils (P < 0.03), lymphocytes (P < 0.0001), monocytes (P = 0.01) and basophils (P = 0.03; Fig. 2I).
Melphalan causes severe histopathological injury in the small and large intestine
Melphalan caused severe architectural injury in the small and large intestine, characterized by severe villus blunting/atrophy and crypt degeneration at day 4 (Fig. 3). Villus height was significantly decreased in melphalan-treated animals compared to controls in the jejunum (481.1 ± 6.5µ m vs 204.3 ± 20.3µ m, P = 0.001, Fig. 3A) and ileum (284.3 ± 12.7µ m vs 143.8 ± 18.6µ m, P = 0.004, Fig. 3B). Crypt depth was significantly increased in the jejunum at day 4 (141.6 ± 3.2 µm vs 161.0 ± 1.7 µm, P = 0.03, Fig. 3C) and day 7 (127.2 ± 4.3 µm vs 170.9 ± 9.7 µm, P = 0.006, Fig. 3C). In contrast, crypt depth in the ileum was significantly decreased at day 4 (145.2 ± 1.1 µm vs 126.3 ± 3.8 µm, P = 0.04, Fig. 3D) in melphalan-treated rats compared to controls, whilst an increase was observed at day 7 (149.4 ± 5.6µ m vs 199.7 ± 4.1µ m, P = 0.006, Fig. 3D). No significant changes in crypt depth were observed for the colon (Fig. 3E).
Strong correlations were observed between villus height and citrulline for both the jejunum and ileum (R2 = 0.68, R2 = 0.62, respectively, P < 0.0001, Fig. 3F). Significant, but less robust correlations were observed for citrulline and crypt depth for the ileum and colon (R2 = 0.31, P = 0.001; R2 = 0.55, P < 0.0001, respectively, Fig. 3G).
In addition to villus blunting and changes in crypt depth, melphalan caused severe architectural derangement in the small and large intestine, characterized by villus fusion and crypt ablation (Fig. 3H), with evidence of gross colonic pathology seen at day 7 (Fig. 3Ii). Histological analysis of a macroscopically evident colonic perforation showed complete destruction of the mucosa, submucosa and muscularis layer (Fig. 3Iii), with pseudomembrane development and visible inflammatory infiltrate (Fig. 3Iiii). Infiltration of polymorphic nuclear cells (PMNC) was also elevated in the jejunum of melphalan treated rats (Fig. 4).
Melphalan-induced microbiota disruption is characterized by pathogen expansion and SCFA deficits
16S rRNA-gene analysis was performed in fecal samples collected longitudinally (control and melphalan, N = 8 per group) on day 0, 4, 7 and 10. Melphalan induced significant disruption of the fecal microbiota that failed to recover during the 10-day experimental period (Fig. 5). Compositionally, there was a shift towards a Firmicute-dominated microbiota, with expansion of pathogenic taxa largely belonging to the Proteobacteria phylum (Fig. 5A) which was not observed in control animals. This was accompanied by a decrease in the number of operational taxonomic units (OTUs), an indicator of microbial richness, which was significantly decreased 7 days post-melphalan compared to controls (P = 0.009, Fig. 5B). There were no significant changes in alpha diversity parameters, including Chao1, Simpson index and Shannon’s index in both control and melphalan-treated rats (data not shown). Principle component analyses showed no change in beta diversity of control animals throughout the experimental period (Fig. 5C). In contrast, significant changes were evident at all time points in melphalan-treated animals compared to baseline (P < 0.0001, Fig. 5D). These changes were significant compared to controls at day 4 (P = 0.021) and day 7 (P = 0.015).
While beta diversity and richness were most profoundly affected at day 7, expansion of pathogens was most significant at day 4 post-melphalan (Fig. 5E), with significant increases in the abundance of Proteobacteria (P = 0.027), Betaproteobacteria (P = 0.021), Gammaproteobacteria (P = 0.015), Burkholderiales (P = 0.015), Enterobacteriales (P = 0.021), Bacteroidaceae (P = 0.015), Enterobacteriaceae (P = 0.018), Bacteroides (P = 0.018), Escherichia-shigella (P = 0.015), Clostridium sp. (P = 0.001), Bacteroides vulgatus (P = 0.006) and Escherichia coli (P = 0.015). This was accompanied by a detectable decrease in Muribaculum (S24-7) in melphalan-treated rats compared to controls (P = 0.017).
Significant decreases in acetate (9.25 ± 0.63 mM vs 3.75 ± 0.75 mM, P = 0.005, Fig. 6A), propionate (35.12 ± 4.02 mM vs 14.75 ± 2.05 mM, P = 0.025, Fig. 6B) and butyrate (5.00 ± 0.58 mM vs 1.75 ± 0.47 mM, P = 0.015, Fig. 6C) were observed at day 4. Significant deficits in propionate and butyrate were also observed at day 10 (P = 0.027, P = 0.021, respectively).
Orally-administered FITC-dextran was used to assess epithelial barrier permeability at all terminal time points. While no significant changes were observed, the highest concentration of plasma FITC-dextran was observed on day 7 post-melphalan treatment (Figure S2). At this time point we also identified one case of positive blood culture (Escherichia coli). All other blood cultures were negative.
Melphalan-induced ileal injury results in bile acid malabsorption and decreased plasma primary to secondary bile acid ratios
Plasma bile acids profiles were analyzed at termination on day 4, 7 and 10. Total plasma bile acid concentrations and primary/secondary ratios were decreased in melphalan treated animals on day 4 and 7 but this did not reach statistical significance (Fig. 7A-B). Upon analyzing individual bile acid species, the primary conjugated bile acid, taurocholic acid (TCA), was decreased at all evaluated time points post-melphalan treatment (Fig. 7C, P = 0.006, P = 0.0004, P = 0.0001, respectively). The rodent specific primary bile acid tauro-alpha-muricholic acid (T-α-MCA) was also decreased at all time points (Fig. 7D, P = 0.01, P = 0.003, P = 0.009, respectively), while beta-muricholic acid was unchanged (Fig. 7E). Secondary bile acid deoxycholic acid (DCA) was unchanged by melphalan treatment in its unconjugated form (supplementary data) but was higher as taurine conjugate at day 4 (P = 0.001, Fig. 7F). All other bile acid concentrations are listed in Table S2.
Ileal gene expression analysis was performed at day 4 for target genes involved in bile acid absorption and transport (Fig. 7G). Ileal expression of t-Asbt, Fabp6 and Ostβ was significantly lower in melphalan treatment compared to controls, suggesting lower ileal absorption of bile acids (P = 0.03). In contrast, farnesoid X receptor (FXR) target genes Shp and Fgf15 were unchanged and increased (P = 0.03), respectively.
{kind=link}