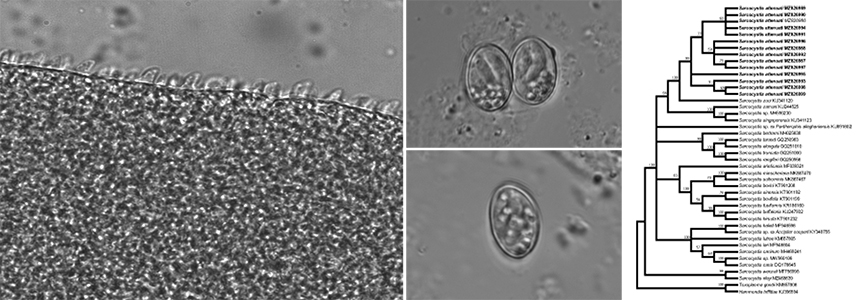
LM and TEM observations of sarcocysts
Spindle-shaped sarcocysts were found in 17 of 42 (40.5%) Asian gray shrews, located in skeletal muscle, the esophagus, the diaphragm, the tongue and the heart. Only one form of sarcocysts was observed. The observation of fresh samples at the light microscopy level showed that the cyst walls of the sarcocysts had numerous, 3.3–4.5 μm-long (n=25) saw-tooth-like protrusions (Fig. 1a). Mature sarcocysts were 740–1,355 × 117–250 μm in size; they were septate and contained bradyzoites measuring 8.2–10.4 × 2.0–3.0 μm (n=40) in size.
Ultrastructurally, the sarcocysts had lancet- or leaf-like villous protrusions measuring 2.8–6.4 × 0.4–0.9 μm (n=15), which contained numerous electron-dense granules in the core; microtubules or fibrils were absent. The primary cyst wall had minute undulations over the entire sarcocyst surface, lined by an electron-dense layer. The nearly vertical protrusions were separated from each other at inconsistent distances. A layer of ground substance measuring 1.2–1.6 μm (n=15) in thickness was located immediately beneath the primary sarcocyst wall (Fig. 1b).
Infection of the definitive host
The two beauty rat snakes fed muscle tissue containing sarcocysts in an Asian gray shrew excreted sporulated oocysts/sporocysts (Fig. 1c, d) in their feces, one beginning on Day 10 and another on Day 11 PI. Upon the death of the snakes at Day 29 PI, numerous oocysts/sporocysts were also observed within the small intestines of the experimental animals. Under light microscopy, the sporulated oocysts measured 11.9–16.7 × 9.2–10.6 μm (average 13.5×9.9 μm) (n = 27), with two elliptical sporocysts measuring 9.2–10.6 × 6.3–6.8 μm (average 9.9 × 6.6 μm) (n = 30). No oocysts/sporocysts were found in the feces or small intestine of the control snake.
Molecular analysis
Seven genes (18S rDNA, ITS1, cox1, cox3, cytb, rpoB and clpC) were successfully amplified from ten individual sarcocysts and oocysts/sporocysts. A total of 16 clones, including ten from each sarcocyst and six from oocysts/sporocysts, were sequenced, assembled, and submitted to GenBank. The ten 18S rDNA sequences obtained from the sarcocysts were 1,863–1,867 bp in length and shared 99.7–100% identity (average 99.8%). The six 18S rDNA sequences obtained from the oocysts/sporocysts were 1,865–1,868 bp in length and shared 99.7–100% identity (average 99.8%). Therefore, only three sequences from sarcocysts (accession numbers MZ826981–MZ826983) and two sequences from oocysts/sporocysts (MZ826984 and MZ826985) were deposited in GenBank. The sequence identity between the sarcocysts and oocysts/sporocysts was 99.7–100%, with an average of 99.8%. The most similar sequences in GenBank were those of S. scandentiborneensis (MN733816 and MN733817) obtained from the tree shrews Tuaia minor and T. tana (ranging 97.6–98.3% identity, on average 97.9%), followed by S. zuoi (ranging 96.7–98.0% identity, average 97.2%) from the Malayan field rat, Rattus tiomanicus (KU341118–KU341121); Sarcocystis sp. (AB251613) from the raccoon Procyon lotor (ranging from 95.9–96.9% identity, average 96.3%); and S. clethrionomyelaphis (KP057504, KF309700, and KF309701) from the large oriental vole, Eothenomys miletus (ranging from 95.9–96.4% identity, average 96.2%).
The ten ITS1 sequences obtained from sarcocysts were 874–978 bp in length and shared 97.9–100% identity (average 99.0%). The six ITS1 sequences obtained from oocysts/sporocysts were 875–877 bp in length and shared 98.6–100% identity (average 99.0%). Therefore, only 14 sequences, including nine from sarcocysts (MZ826986–MZ826994) and five (MZ826995–MZ826999) from oocysts/sporocysts, were deposited in GenBank. The similarity between the sarcocysts and oocysts/sporocysts was 97.4–100%, with an average of 98.7%. The most similar sequences in GenBank were those of S. zuoi (KU341118–KU341121) from the Malayan field rat, but the identity was only 72.4–86.0% (average 76.4%).
The ten cox1 sequences obtained from sarcocysts were 1,333 bp in length and shared 99.8–100% identity (average 99.9%). The six cox1 sequences obtained from oocysts/sporocysts were 1333 bp in length and shared 99.9–100% identity (average 99.9%). Therefore, only five cox1 sequences, including three from sarcocysts (MZ889669–MZ889671) and two from oocysts/sporocysts (MZ889672 and MZ889673), were deposited in GenBank. The similarity between the sarcocysts and the oocysts/sporocysts was 99.8–100%, with an average of 99.9%. The most similar sequences in GenBank were those of S. scandentiborneensis (MN732561 and MN732562, 100% identity), followed by those of Sarcocystis sp. (MT411016, 99.8% identity) from the greater white-toothed shrew, Crocidura russula; and S. canis (KX721496 and KX721497) from the Indo-Pacific bottlenose dolphin, Tursiops aduncus (ranging 95.0–95.4% identity, average 95.2%).
The 16 cox3 sequences from the sarcocysts and the oocysts/sporocysts were 675 bp in length and were completely identical. Therefore, only one sarcocyst sequence (OK001462) and one oocyst/sporocyst sequence (OK001463) were deposited in GenBank. No sequences with significant similarity to these sequences were found in GenBank.
The 16 cytb sequences from the sarcocysts and the oocysts/sporocysts were 1,080 bp in length and were completely identical. Therefore, only one sarcocyst sequence (OK001464) and one oocyst/sporocyst sequence (OK001465) were deposited in GenBank. The most similar sequences in GenBank were those of S. falcatula (MF034168–MF034187) from the budgerigar, Melopstittacus undulatus, and the identity was 96.4%.
The 16 rpoB sequences obtained from the sarcocysts and the oocyst/sporocysts were 511 bp in length and shared 100% identity. Therefore, only one sarcocyst sequence (OK001466) and one oocysts/sporocysts sequence (OK001467) were deposited in GenBank. The most similar sequence in GenBank was that of S. neurona (GQ851961) obtained from the southern sea otter, Lutris nereis, and the identity was 91.9%.
The 16 clpC sequences from the sarcocysts and the oocysts/sporocysts were 534 bp in length and shared 100% identity. Therefore, only one sarcocyst sequence (OK001468) and one oocyst/sporocyst sequence (OK001469) were deposited in GenBank. The most similar sequences were those of S. falcatula (KP871717) and S. neurona (KP871716), and the identity was 92.5%.
Phylogenetic analysis
Phylogenetic analysis based on 18S rDNA, ITS1, or cox1 sequences confirmed that the parasite found in the present study belonged to Sarcocystis (Figs. 2–4). In the phylogenetic tree inferred from 18S rDNA sequences (Fig. 2), the parasite formed an individual clade and clustered with Sarcocystis spp. using colubrids as definitive or presumed definitive hosts (i.e., S. scandentiborneensis (MN733816), S. zuoi (KU341120), S. clethrionomyelaphis (KP057504) and Sarcocystis sp. ex Procyon lotor (AB251613)). These species belonged to a group containing Sarcocystis spp. with pythons as definitive hosts (i.e., Sarcocystis sp. ex Morelia viridis (KC201639), S. zamani (KU244524) and S. singaporensis (AF434054)). In the phylogenetic tree inferred from ITS1 sequences, the parasite formed an individual group and clustered with Sarcpcystis spp. with sankes as definitive or presumed definitive hosts (i.e., S. zuoi (KU341120), S. zamani (KU244525), Sarcocystis sp. (MH590230) and S. singaporensis (KU341123)). In the phylogenetic tree inferred from cox1 sequences (Fig. 4), the parasite formed an individual clade with S. scandentiborneensis (MN732562).
Based on the morphological characteristics of sarcocysts, molecular analysis and host specificity, a new species name, Sarcocystis attenuati, is proposed for the organism found in Asian gray shrews from Anning Prefecture, China.
Taxonomic summary of Sarcocystis attenuati n. sp.
Diagnosis: The sarcocysts were microscopic, 740–1,355 μm long, and 117–250 μm wide. The thick-walled sarcocysts had 3.3–4.5 μm-long saw-tooth-like protrusions. The sarcocysts were divided by septa into a series of internal compartments filled with bradyzoites measuring 8.2–10.4 × 2.0–3.0 μm. TEM revealed sarcocysts with lancet- or leaf-like protrusions measuring 2.8–7.4 × 0.4–0.9 μm (n=15), which contained electron-dense granules in the core. The nearly vertical protrusions were separated from each other at inconsistent distances, similar to wall type 9h classified by Dubey et al. (2016).
Type (natural) intermediate host: Asian gray shrew, Crocidura attenuata.
Type locality: Anning Prefecture (25°24′N, 102°35′E; altitude 1,894 m), China.
Site of infection: Muscular tissues throughout the body, including the heart.
Experimental definitive host: beauty rat snake, Elaphe taeniura.
Prevalence: Sarcocysts were found in 17 of 42 (40.5%) examined Asian gray shrews.
Etymology: Species named after the species of its intermediate host.
Molecular characterization: Nucleotide sequences of the 18S rDNA (MZ826981–MZ826985), ITS1 (MZ826986–MZ826999), cox1 (MZ889669–MZ889673), cox3 (OK001462 and OK001463), cytb (OK001464 and OK001465), rpoB (OK001466 and OK001467), and clpC (OK001468 and OK001469) of the new species have been deposited in GenBank. According to 18S rDNA sequences, S. attenuati may be differentiated from S. scandentiborneensis obtained from lesser or large tree shrews. However, the cox1 sequences are unsuitable for differentiating it from S. scandentiborneensis or even Sarcocystis sp. obtained from the greater white-toothed shrew.
Specimens deposited: Formalin-fixed tissues containing cysts of S. attenuati as well as photomicrographs obtained from the LM and TEM examination of the sarcocysts were deposited at the Zoological Specimen Museum of Yunnan University, Kunming, China (collection number Pro2018004).
Remarks
Insectivores are an abundant group of small mammals comprising hedgehogs, moonrats, shrews, and moles. Publications regarding Sarcocystis spp. in insectivores are very limited. To our knowledge, sarcocysts have been observed only in several species of insectivores, including the shrew mole Urotrichus talpoides in Japan [11], moonrat, Echinosorex gymnurus, in Malaysia [2], the short-tailed shrew Blarina brevicauda in the USA [12], the white-toothed shrew Crocidura russula in Russia [3], and the common shrew Sorex araneus in Lituanica [13]. Unfortunately, the descriptions of most sarcocysts recorded in the abovementioned cases are not detailed. Based on the morphological characterization of sarcocysts, only two species, S. boollati and S. russuli, have been proposed, which were found in moonrat and white-toothed shrews, respectively. With the aid of LM, it was shown that both species exhibit smooth, thin sarcocyst walls (< 1 μm) [2, 3]. Ultrastructurally, S. booliati sarcocysts show small knob-like protrusions, similar to wall type 1b [14]. In the present study, S. attenuati n. sp. found in Asian gray shrews presented a thick sarcocyst wall (3.3–4.5 μm in length), similar to wall type 9h, which was unambiguously different from those of S. boollati and S. russuli.
The ultrastructure of sarcocysts has traditionally been used as a reliable indicator for the characterization of different Sarcocystis species in a given host. Nucleotide sequence analysis has now been suggested to be a more useful tool for the delineation or identification of Sarcocystis from the same or different hosts. However, different genetic markers have revealed different levels of intra- or interspecific sequence diversity [15–17]. Here, seven molecular markers (18S rDNA, ITS1, cox1, cox3, cytb, rpoB, and clpC) were sequenced and characterized in DNA samples from S. attenuati. The analysis showed that the sequences of these seven genes of the parasite presented high intraspecific similarities (i.e., 99.7–100%, 97.9–100%, 99.8–100%, 100%, and 100%, and 100%, 100%, respectively). The comparison of these sequences with those previously deposited in GenBank showed that the 18S rDNA and cox1 sequences of S. attenuati shared the highest identity with those of S. scandentiborneensis (i.e., 97.6–98.3% and 100%, respectively). Sarcocystis scandentiborneensis occurs in lesser or large tree shrews belonging to the family Tupaiidae collected from Malaysia. With the aid of LM, the cyst wall of S. scandentiborneensis was shown to present tightly packed figure-like protrusions (2–10 μm in length) that can assume a brush-like appearance; ultrastructurally, the tightly packed figure-like protrusions contain bundled microtubules that extend into the ground substance [18], similar to wall type 11b or 12 according to the classification of Dubey et al. (2016) [1]. Additionally, the cox1 sequences of S. attenuata shared a high similarity of 99.8% with that of Sarcocystis sp. obtained from the greater white-toothed shrew collected from Spain. However, sarcocysts of the greater white-toothed shrew have not been described [19].
The phylogenetic relationships among the majority of analyzed Sarcocystis spp. suggest their coevolution with their definitive hosts rather than their intermediate hosts [20]. The phylogenetic trees based on 18S rDNA sequences and ITS1 sequences revealed that S. attenuati formed an individual clade with Sarcocystis spp. from small mammals using snakes as definitive or putative definitive hosts, showing especially close relationships with S. scandentiborneensis, Sarcocystis sp. ex Procyon lotor, S. zuoi, and S. clethrionomyelaphis.
The infection experiment confirmed the speculation arising from the phylogenetic analysis and proved that beauty rat snakes can serve as an experimental definitive host of S. attenuati. The beauty rat snake is native to eastern and southeastern Asia and feeds mainly on shrews and rodents, although the consumption of amphibians, reptiles, and birds by this species has also been reported [21]. To date, only two species of Sarcocystis, S. zuoi from Norway rat, R. norvegicus, and S. clethrionomyelaphis from large oriental vole, E. miletus, have been proven to use species of Elaphe as definitive hosts via transmission experiments [22, 23]. Morphologically, sarcocysts of S. zuoi exhibit sloping finger-like protrusions, and the base of the protrusions is highly branched [22, 24], similar to wall type 17; the sarcocysts of S. clethrionomyelaphis present thin highly folded protrusions, which often bend along the cyst surface [23], similar to type wall type 10f. Therefore, the sarcocysts of S. attenuati can be easily morphologically differentiated from those of S. zuoi and S. clethrionomyelaphis. It is very common that one predator acts as the definitive host of more than one species of Sarcocystis. For example, Python reticulatus is the definitive host of three species of Sarcocystis in rats (i.e., S. singaporensis, S. villivillosi, and S. zamani) [25].
{kind=link}