Overview of the Genomic metadata Composition of K. pneumoniae isolates
The PATRIC metadata depicts 144 clinical strains and 9 environmental strains. The isolation source of strains are blood (n=115), urine (n=9), endotracheal aspirate (n=5) bronchoalveolar lavage (n=4), sputum (n=4), pus (n=3), CSF (n=2), nasal swab (n=2), rhizosphere soil (n=1; AWD5), root nodule (n=1; HPCN22), domestic sewage (n=2; HPCN5, HPCN17), STP sludge (n=1; PVN-1), endophyte (n=1; ME30), river surface water (n=1; PL1-RCS238), industrial waste water (n=1; EGD-HP19), agriculture field soil (n=1; KBG6.2) (Supp.table 1). The genomic characterization of K. pneumoniae was performed as first part of the study. From the PARTIC database, we found these genomes were sequenced majorly by illumina and ion torrent. The genomes of study strains ranged in size from 4.4 Mbp to 6.3 Mbp and GC contents were between 55-57%.
Sequence type
The study isolates were found to have 37 different STs, suggesting its diversity in India isolates. We found ST231 was predominant (n=43) among clinical strains. Followed by, ST147 (n=15), ST14 (n=15), ST2096 (n=11), ST395 (n=9), ST43 (n=8), ST 11 (n=7), ST23(n=6), ST16 (n=6), ST15 (n=3), ST437 (n=2), other clinical strains comprised individual STs, ST13, ST42, ST101, ST307, ST557, ST570, ST628, ST3249, ST711, ST660, ST3836, ST3835, ST3789, ST3607, ST3605, ST4847, ST2816, ST3249. The scatter plot represents the distribution of STs, where ST231, ST147, ST2096 emerged and displayed with increasing trends during 2013, 2015 and 2018 (Figure 1). Among these, ST14 was common across all years. The environmental strains comprised of individual STs namely, ST22, ST200, ST555, ST1107, ST1728, ST2701, ST3689.
Phylogenetic analysis
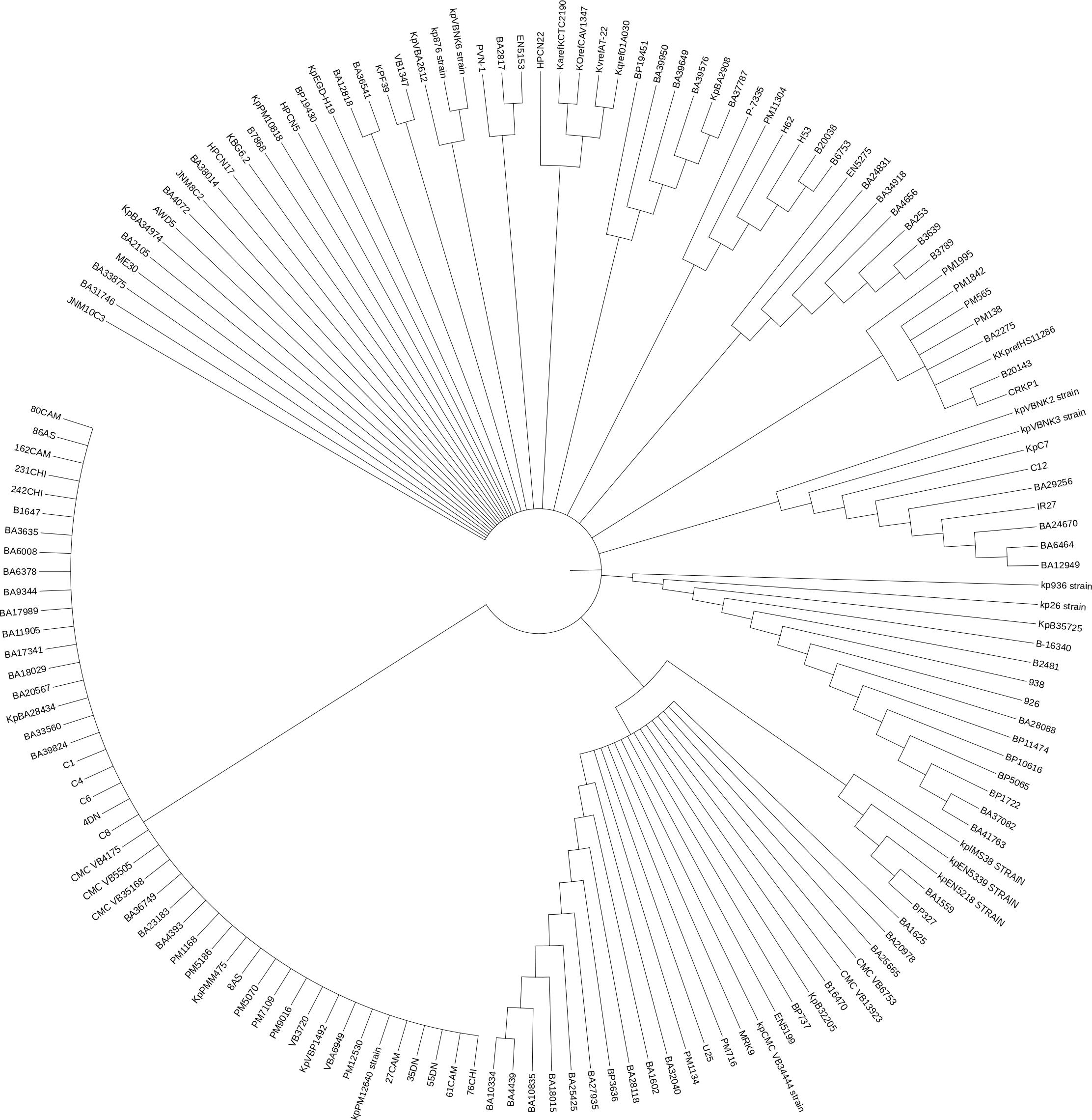
The WGS of study strains were used for the phylogenetic analysis based on MLST (Suppl. Figure 1) and Single nucleotide polymorphism (Figure 2). The midpoint was divided into 2 major clades and further delineated into many clusters of K. pneumoniae. The environmental strain HPCN5, HPCN17, HPCN22, PL1-RCS238, AWD5, KBG6.2, EGD-HP19 were grouped in clade 1, whereas ME30 the endophyte isolate aligned in clade 2, suggesting the evolution as a separate ancestral group. The reference strain MGH78578 was closely aligned with environmental strain PVN1, STP sludge isolate. The environmental strains were identified on a separate branch of the clusters consisting of clinical strains.
K. pneumoniae has a high prevalence of β-lactam and fluoroquinolone resistance
Using the ResFinder and CARD databases, 88 different predominant AMR genes involved in 8 antibiotic classes were selected for AMR profiling. The resistance was found as follows, fosfomycin 100% (n=153), β-lactam 96% (n=147), fluoroquinolone 92% (n=142) followed by aminoglycoside 84% (n=129), sulfonamide 79.08% (n=121), phenicol 70 % (n=107), macrolide 64.04% (n=98), rifamycin 48 % (n=74). AMR genes associated with fosfomycin (FosA-like), β-lactam (blaCTX-M-15, blaTEM-1B), fluoroquinolone (oqxA-like, oqXB-like) aminoglycoside (aadA2, aac(6') lb-cr-like) were predominant across the India strains (Suppl. Table 2).
To understand the distribution of AMR genes in 153 genomes during 2010-2020, Box and whisker plot was used (Figure 3). The data revealed the presence of a genome having a least of one (n=1) and a maximum of 26 (n=4) AMR genes. The average median distributions of 15 AMR genes/genome were noticed during 2013-2017.
Most commonly, β-lactamase genes blaCTX-M-15 (n=92), blaTEM-1B (n=87), carbapenamases blaOXA-232 (n=66), metallo-β-lactamase blaNDM-5 (n=29) and blaNDM-1(n=10) were found among the study strains. blaNDM-5 exhibited co-positivity with blaCTX-M-15 and blaTEM-1B. The strains carrying blaCTX-M-15, blaTEM-1B, and blaNDM-5 were increased from 2019.
Followed by, drug resistance mechanisms involved in study strains depicts antibiotic efflux (57.51%, n=88) and antibiotic inactivation by hydrolytic enzymes (41%, n=62) as predominant mechanisms. The major gene family observed for antibiotics efflux are resistance nodulation cell division (RND) (23 %, n=35), major facilitator superfamily (MFS) (18.95%, n=29) and ATP-binding cassettes (ABC) (15.68%, n=24). Similarly, in case of antibiotic inactivation, SHV β-lactamase gene family was prevalent (26.79% n=41) followed by, OXA- β-lactamase (6.9%, n=10) and CTX-M- β-lactamase (3.47%, n=5).
The predominant AMR gene in each 8 class of antibiotics was identified and screened its prevalence among 153 genomes. Such as, blaCTX-M-15 and blaTEM-1B (β-lactam) aadA2 (Aminoglycoside) oqx-A-like and oqx-B-like (Fluoroquinolone) FosA-like: (Fosfomycin), erm(B)-like (Macrolide), sul1 (Sulfonamide), ARR-2 (Rifamycin), catA1like (Phenicol). About 9.8 % of clinical isolates (n=15) were found to have all these 8 listed AMR genes, which are mostly belong to ST231.
The environmental strains also carried the blaSHV-1, FosA-like, efflux pump mediated resistance (oqx-A like, oqx-B-like). Notably, the industrial effluent isolate carried FosA-like, oqx-A-like, oqx-B-like, sul1, aac(6)lb-cr-like, blaSHV112, ere (A)-like, aacA4-like, aadA1.
IncF-type conjugative plasmid is dominant among Klebsiella
To understand the transmission of AMR determinants through plasmids, the type of plasmid and the resistance genes they carry were investigated. From the metadata, the number of plasmids estimated to be ranged from 1- 8 plasmids per strain, with most isolates harboring at least 2 plasmids. A clinical strain KpIMS38 isolated from pus harbored 16 plasmids. Followed by, the episome details of 153 strains were obtained using Plasmid finder. A total of 19 plasmid replicon types were detected from 93.4 % (n=143) of isolates. The 19 plasmid replicon types are categorized as incompatibility plasmids (Inc) and mobilizable colicin plasmid (Col) groups. The most abundant epidemic plasmid is IncF-type conjugative plasmid (80.39 % n=123) (Figure 5). Plasmids belonging to IncF-type are reported to carry ESBL genes, genes encoding carbapenemases, aminoglycoside modifying enzymes and quinolone resistance genes (Rozwandowicz et al., 2018). The colKP3 plasmid was identified in 83 isolates which is related to blaOXA-232 and blaOXA-181 transfer. The second most prevalent were pKpQIL-IT plasmids (n=75) belonging to IncFIB (pQIL) family carries an aminoglycoside resistance gene on a transposon-like element IS26. Both colKP3 and pKpQIL-IT plasmids were accompanied in 33.9% (n=52) isolates. These two dominant plasmids were not detected in any of the environmental strains. IncHI1B (pNDM-MAR) plasmid with NDM resistance was found in 33 clinical strains and an environmental strain. The colistin resistance genes (mcr1) encoding plasmids (IncX4, IncY) were detected only in three strains among 153 study isolates. Certain resistant genes blaCTX-M-15 (IncR, IncFII, IncFII, IncHI1B, Col440I), blaNDM-5 (IncFII, IncX3) were detected in various plasmid replicon types. In the case of environmental strains, 5 isolates were harbored with one plasmid. The endophyte isolate has 2 plasmids possessing blaCTX-M-15, blaNDM, qnrB1, blaSHV-56, FosA, oqxA and oqxB resistant genes. The industrial effluent isolate has IncY, Phage-like plasmids encoding mcr-1 gene, we have also recorded the multiple resistance being carried by this isolate. In summary, clinical and environmental strains have plasmids encoding genes that confer resistance to β-lactam drugs, aminoglycoside, quinolone and colistin. The type, family and functions of plasmids are provided in Table 1.
Correlation of Resistance and Plasmid type
We narrowed down to identify the strains that carry predominant AMR genes among the 8 selected antibiotic classes (n=15). These clinical isolates were found to have pKpQIL-IT and pKP3-A plasmid types. pKpQIL-IT carries aminoglycoside resistance gene on a transposon-like element IS26, a clinically important insertion sequence. pKP3-A, a mobilizable but non-self-conjugative plasmid is associated with bla-OXA-232 and blaOXA-181 genes.
Associated virulence genes are higher in ST231 strains
The presence of different virulence genes is shown in (Figure 4), which were screened using VFDB. Both clinical and environmental isolates were found to have genes involved in type1 fimbriae (fimD, fimK, fimH, fimC). Similarly, most isolates were detected with mrkD, mrkJ, mrkF, mrkC, mrkI, mrkA which are important genes for type 3 fimbriae. To identify the hypervirulent phenotypes, rmpA a gene regulator for mucoid phenotype was analyzed. Nearly, 47% (n=77) of clinical and 33.3% (n=3) of environmental strains were identified as hypervirulent strains. Besides, the prevalence of magA gene (mucoviscocity associated gene) was comparatively less in clinical strains (n=56, 38.8%) than the rmpA genes. The magA was detected in environmental strains (n=5, 55.5%) as well. The presence of both rmpA and magA genes was not detected in root nodule, STP sludge and agriculture field soil isolate. The co-occurrence of rmpA and magA genes was observed in domestic sewage isolates (n=2) and an industrial wastewater isolate (n=1). Likewise, the rmpA and magA were accompanied in clinical strains (n=38). These clinical strains were identified as blood isolates majorly with K51 serotype (n=18). Among the study strains, K51 (n=43) and K64 (n=33) serotypes were majorly found. Yet, the K51 serotype was not associated with severe infections and a recent report has identified K64 belonging to ST11 as an emerging superbug worldwide (Zhao et al., 2020), however, none of the 153 Indian isolates were found to have K64-ST11.
The most reported hypervirulent K1 and K2 serotypes (Brisse et al., 2013) were less prevalent, n=7 and n=11 respectively. The co-occurrence of rmpA and magA genes was observed in K1 (n=6) and K2 (n=2) serotypes belong to ST23 and ST14 respectively.
Followed by, most important siderophores were screened. The three most important siderophores systems in Enterobacteriaceae are yersinibactin (ybtS), aerobactin (iutA) and enterobactin (entB). Hence the prevalence of these siderophore systems was analysed in K. pneumoniae strains. The entB was detected in all the study strains (n= 153), ybtS gene was found in 98.6% in clinical (n=142) and 77.7 % in environmental strains (n=7) and iutA gene were observed in 97.2 % in clinical and 88.8% of environmental strains. The root nodule strain was negative for both ybtS and iutA. In addition, allantoin utilization (allS) was screened in the study strains as it has been associated with hypervirulence (Shon et al., 2013). The allS gene was observed in 65.97% of clinical (n=95) and 77.77 % of environmental strains (n=7). Further, the strains associated with hypervirulence factors (magA, rmpA, allS, entB, ybtS, iutA), were identified in 3 environmental strains (EGD HP-19, HPCN17 and HPCN5) and 29 clinical strains. The number of associated virulence genes appeared to be high in strains ST231- K51 and ST23- KL1.
Convergence of AMR and Virulence
To investigate the relationship between AMR and virulence in K. pneumoniae, ESBL (blaCTX-M-15, blaTEM-1B) and carbapenamase (blaOXA-232) harboring isolates were mapped with major virulence genes (rmpA, magA, ybtS, alls, iutA). 98% of the ESBL and 62% of carbapenamase isolates harbored ybtS, iutA and rmpA, magA respectively. The positive correlation between resistance and virulence in K. pneumoniae seeks prominent attention in clinical settings.
{kind=link}