EPB41L4A-AS1 is down-regulated in aging and neurodegenerative diseases
EPB41L4A-AS1 is a metabolism-related lncRNA. Brain metabolic patterns are altered in aging and neurodegenerative diseases, especially in AD. Therefore, we think the expression of EPB41L4A-AS1 may be dysregulated in these conditions. To address these issues, the Mann-Whitney test method was used to compare the expression of EPB41L4A-AS1 in different age groups, and between normal and AD groups. As shown in Fig. 1A-B, EPB41L4A-AS1 expression gradually decreases with age increasing, and its expression is also down-regulated in the AD group, compared with normal people.
Differences of gene expression in the hippocampus of different age groups
The hippocampus is the main damaged area in aging and neurodegenerative diseases, so gene expression profiles (32972 genes) of 197 samples from the hippocampus (31 samples 20-50 years old, 52 samples 50-60 years old, 114 samples 60-80 years old) were used for gene differential expression analysis (age-related). To get better results, the expression profiles of two sets of samples (20-50 years old, 60-80 years old) were used as an expression matrix.
A total of 6538 DEGs (3246 up-regulated and 3292 down-regulated) between the elderly group (60-80 years) and the young group (20-50 years) were screened out under the threshold of p-Value < 0.05. The down-regulation of EPB41L4A-AS1 is also confirmed in the volcano plot of DEGs (Fig. 1C).
Construction of Co-Expression Modules
To find genes having similar expression patterns with EPB41L4A-AS1, the expression profile of all DEGs in 197 samples from the hippocampus was extracted to construct a weighted co-expression network using the WGCNA algorithm. The appropriate samples were screened out by sample clustering (Fig. 1D), and the appropriate power value was screened out (Fig. 1E-F). Scale-free networks were constructed with proper independence degree and average connectivity when the power value was equal to 14. The DEGs with similar expression patterns were cluster into four distinct gene co-expression modules (Fig. 1G).
Interaction Analysis with Clinical Traits and GO, KEGG Enrichment
The heatmap (Fig. 2A) demonstrates the Topological Overlap Matrix (TOM) among 400 randomly chosen genes in the analysis. The correlation of three modules successfully clustered is shown in the heatmap (Fig. 2B). As a result, each module is independent of the other.
EPB4L4A-AS1 is located in the turquoise module, so we speculate that the protein-coding genes in the turquoise module may be regulated by EPB41L4A-AS1. At the same time, module-trait relationships (Fig. 2C) show that there is a negative correlation (r = -0.35, p < 0.001) between the genes in the turquoise module and age.
To obtain the further function of the turquoise module, GO and KEGG enrichments are applied to all the genes in the turquoise module. The top 10 GO terms and KEGG pathways are visualized in Fig. 2D-E. As showed in the figures, genes in the turquoise module are mainly enriched in nerve excitement transmission and neurodegenerative diseases (such as Alzheimer’s disease (AD), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), Parkinson’s disease (PD), and Prion disease) pathway.
Correlation of EPB41L4A-AS1 and Protein-coding Genes (PCGs)
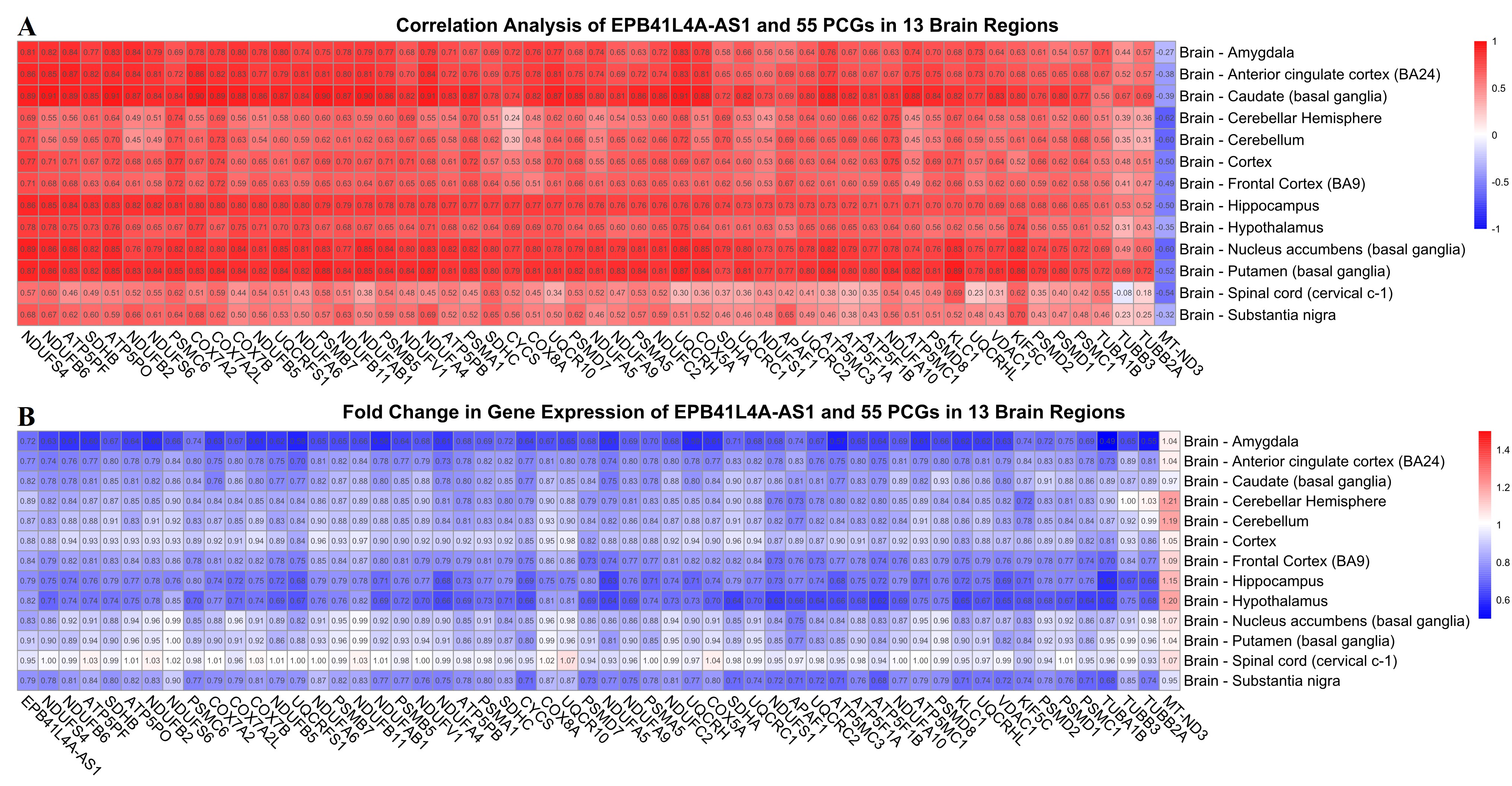
The genes of five neurodegenerative disease-related pathways are processed by the intersection (Fig. 3A). As a result, 55 PCGs are related to all five diseases. A more in-depth GO enrichment analysis of these 55 genes shows that these 55 genes mainly influence oxidative phosphorylation, ATP metabolic process, and electron transport (Fig. 3B). Therefore, we speculated that these 55 PCGs play very important roles in aging and neurodegenerative diseases. It has been proved by cluster analysis that the expression patterns of EPB41L4A-AS1 and these PCGs are similar, thus correlation analysis of EPB41L4A-AS1 and these PCGs is necessary. It shows that EPB41L4A-AS1 is mainly positively correlated with these 55 PCGS in the hippocampus (Fig. 3C) and EPB41L4A-AS1 and these 55 PCGs are mainly down-regulated as brain ages (Fig. 3D). To prove the universality of this relationship, the correlation analysis of 13 brain regions is displayed in the heatmap (Fig. S1A). Furthermore, the fold changes in gene expression of EPB41L4A-AS1 and 55 PCGs in 13 brain regions show that EPB41L4A-AS1 and 55 PCGs are universally down-regulated in the brain with aging (Fig. S1.B). As a result, the relationship is universal.
Gene Function Verification in AD Dataset
To investigate if there are some similarities of the expressive pattern of EPB41L4A-AS1 related genes between the aging group and AD neurodegenerative diseases group, gene expression dataset GSE5281 was used in the next step. For technical reasons, the expression data of 45 PCGs among 55 PCGs were detected in dataset GSE5281. Therefore, we applied the correlation analysis of EPB41L4A-AS1 and 45 PCGs in GSE5281 (Fig. 3E). The result shows that there is a strong positive correlation between EPB41L4A-AS1 and other PCGs. Also, EPB41L4A-AS1 and these PCGs are down-regulated in AD samples (Fig. 3F). The results further prove that the relationship we proposed before is reliable.
Protein-Protein Interaction Network of 55 PCGs
To reveal the protein-protein interaction network of the 55 PCGs, 55 PCGs were uploaded to STRING and the network is shown in Fig. 4A. The 55 PCGs are mainly the components of complex I-V, which mainly affect the third stage of cellular respiration through molecular biological analysis. The function of complex I is to catalyze the production of NAD+ from NADH [24,25] while complex I-V can catalyze the production of ATP from ADP by affecting the electron transport chain [26,27].
At this stage, we found that EPB41L4A-AS1 has a strong positive correlation with 55 PCGs, and these 55 genes were involved in the formation of complex I-V on the electron transport chain. By analyzing the reactions related to the electron transport chain, we speculated that the down-regulation of EPB41L4A-AS1 will down-regulate the expression of the components of complex I-V, thus reducing the activity of complex I-V, and ultimately leading to the down-regulation of NAD+ and ATP levels.
Previous studies have reported that brain NAD+ is associated with ATP energy production and membrane phospholipid turnover in humans [28], and ATP plays a very important role in the normal brain [29]. Furthermore, NAD+ has been reported to be strongly related to aging and neurodegenerative diseases [18,30], and NAD+ can inhibit Alzheimer’s disease [31]. As a result, when NAD+ and ATP levels go down, the body’s energy and anti-aging ability will decrease, which may eventually lead to aging and neurodegenerative diseases [32].
Analysis of NAD+ synthesis pathway genes
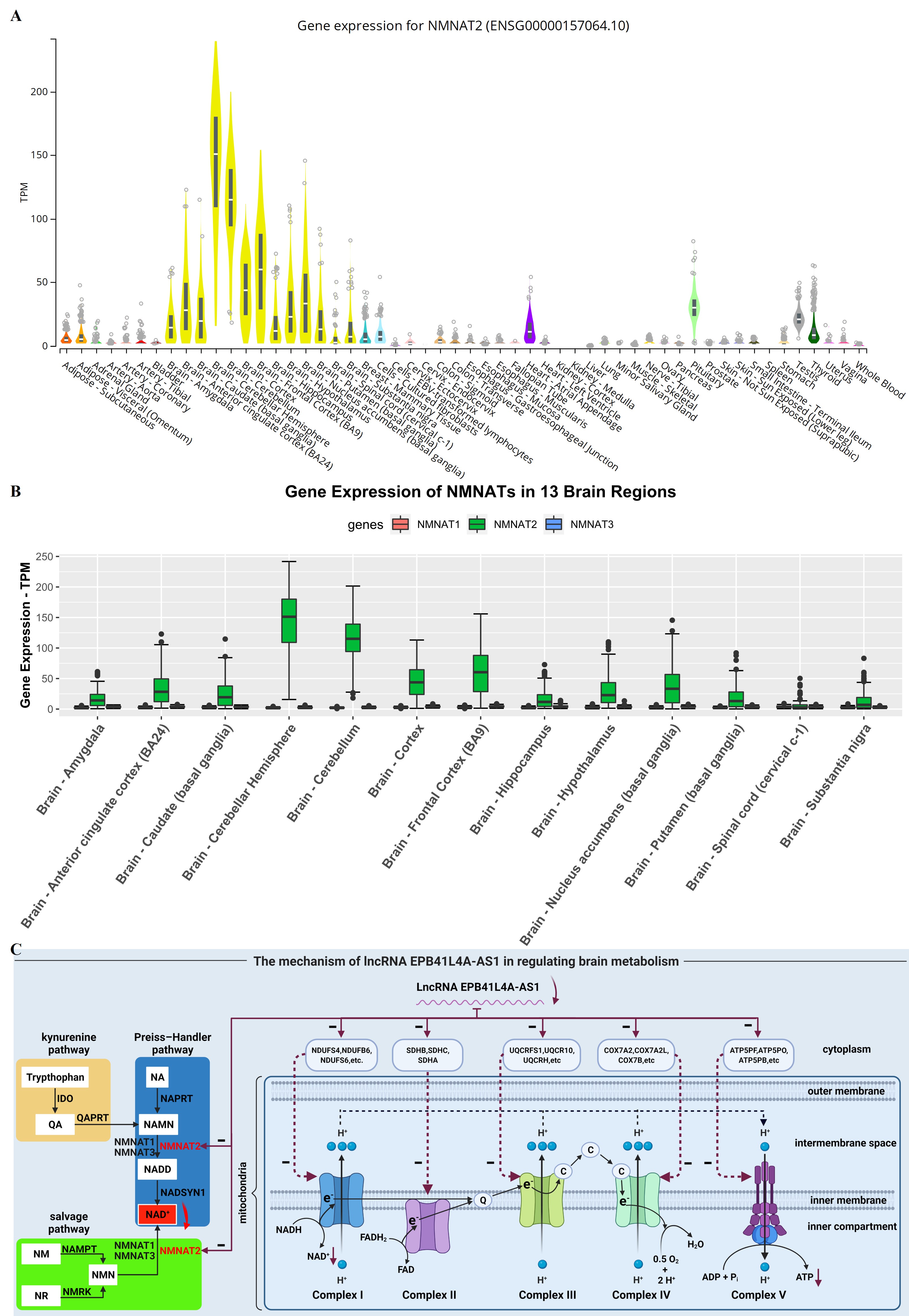
NAD+ level is not only related to electron transport chain but also maintained by three independent biosynthetic pathways, kynurenine pathway (or de novo synthesis pathway), Preiss–Handler pathway, and salvage pathway [16]. Furthermore, it has been reported that the kynurenine pathway is associated with neurodegenerative diseases [33,34], while the salvage pathway protects neurons from chemotherapy-induced degeneration [35]. Inspired by these, we analyzed the expression levels of genes in the NAD+ biosynthesis pathways in aging and neurodegenerative diseases. As a result, we found that only NMNAT2 (enriched in the cytoplasm [36]) is a DEG and it mainly expresses in the brain (Fig. S2A). NMNATs catalyze the reaction of NAMN (Preiss–Handler pathway) and NMN (salvage pathway) to form NAD+ and NMNAT2 is the main form of NMNATs in the brain (Fig. S2B). In addition, NMNAT2 is located in the same module (turquoise module) as EPB41L4A-AS1 in the WGCNA process, and it’s also a down-regulated gene with aging (Fig. 4B). Correlation analysis of EPB41L4A-AS1 and NMNAT2 also showed that EPB41L4A-AS1 has a strong positive regulation on NMNAT2 in aging (Fig. 4C). For further study, we analyzed the expression level of NMNAT2 in normal and AD samples and its correlation with EPB41L4A-AS1 in dataset GSE5281. As shown in Fig. 4D-E, NMNAT2 is down-regulated in Alzheimer’s disease and has a strong correlation with EPB41L4A-AS1.
In short, EPB41L4A-AS1 may regulate the NAD+ and ATP generation via impacting the genes of the electron transport chain and NAD+ synthesis pathway (NMNAT2), and thus plays an important role in brain aging and neurodegenerative diseases. The mechanism is shown in Fig. S2C.
EPB41L4A-AS1 has a positive regulation of NAD+ and ATP synthesis in SH-SYSY and U251 cells
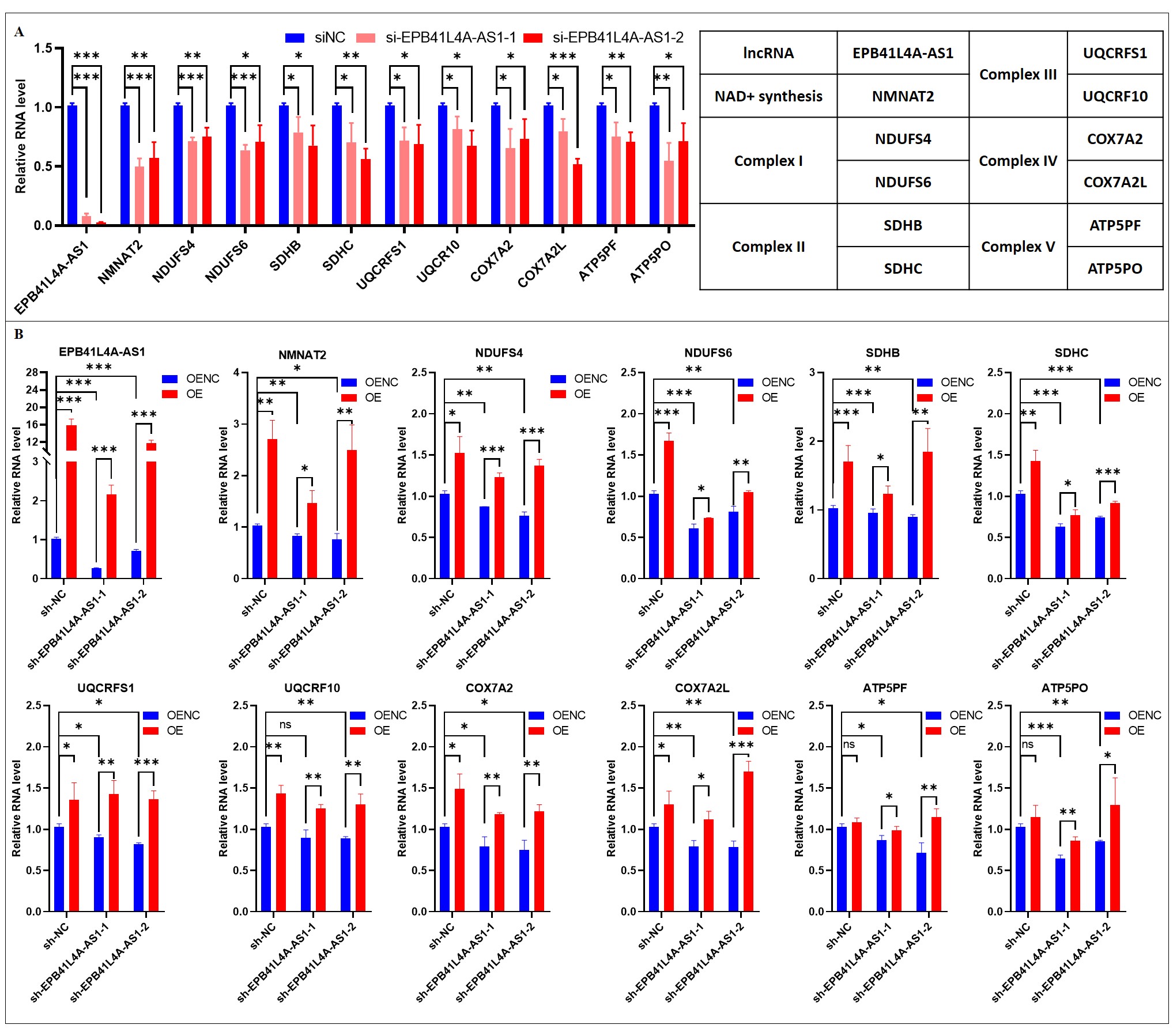
In this process, we applied experiments on SH-SY5Y and U251 cells to prove the role and mechanism of EPB41L4A-AS1 in brain aging and neurodegenerative diseases. For each complex of the mitochondrial respiratory chain, we selected two PCGs with the highest correlation coefficient with EPB41L4A-AS1 for experiments. As a result, a total of 11 PCGs (10 genes of 5 complexes, NMNAT2) were used to check if EPB41L4A-AS1 plays a regulatory role in them.
Firstly, we used siRNAs to knock down EPB41L4A-AS1 in SH-SY5Y cells to observe the changes of 11 PCGs. Two siRNAs (si-EPB41L4A-AS1-1, si-EPB41L4A-AS1-2) were successfully transiently transfected and the relative mRNA expressions of 11 target genes were down-regulated (Fig. 5A). We also applied sh-EPB41L4A-AS1 lentivirus transfection to obtain EPB41L4A-AS1 stable knockdown cell lines in the next step so it can prove if the changes of 11 target genes are stable when EPB41L4A-AS1 is knocked down. Two sh-EPB41L4A-AS1 cell lines (sh-EPB41L4A-AS1-1, sh-EPB41L4A-AS1-2) were obtained and the results are as same as transient transfection (Fig. 5B). Finally, EPB41L4A-AS1 overexpression plasmids were successfully transfected in the sh-NC cell line and two sh-EPB41L4A-AS1 cell lines. As shown in Fig. 5B, the expression levels of the 11 PCGs rebound when EPB41L4A-AS1 is overexpressed.
To increase the reliability of cell experiments, we carried out the same experiments on U251 cells in the next step. As shown in Fig. S3, the expressions of 11 target genes were down-regulated when siRNAs were transiently transfected (Fig. S3A) and sh-EPB41L4A-AS1 lentivirus transfections were applied (Fig. S3B). Also, the expressions of the 11 target genes were up-regulated as EPB41L4A-AS1 is overexpressed (Fig. S3B).
In the next step, we applied Western blot analysis to reveal the protein levels of the NAD+ and ATP synthesis genes in SH-SY5Y and U251 cells. For ATP synthesis genes, NDUFS4 and NDUFS6 were chosen as the representative genes because the proportion of genes constituting complex I is the largest in previous bioinformatics analysis. Therefore, the protein levels of NMNAT2, NDUFS4, and NDUFS6 were measured in this step. As shown in Fig. 6A and Fig. 6B, the protein levels of these genes are down-regulated when EPB41L4A-AS1 is knocked down, and once EPB41L4A-AS1 is overexpressed, their protein levels were up-regulated.
We finally measured NAD+, NADH, and ATP levels in SH-SY5Y and U251 cells. As shown in Fig. 6C and Fig. 6D, NADH is not changed significantly while NAD+, NAD+/NADH ratio, and ATP levels are down-regulated in sh-EPB41L4A-AS1 cells, and overexpression of EPB41L4A-AS1 upregulated them. So far, we have proved the correctness of our proposed regulatory mechanism from the perspectives of bioinformatics, molecular biology, and cell biology.
NR treatment can improve the NAD+ and ATP levels in SH-SY5Y and U251 cells
Overexpression of EPB41L4A-AS1 can restore the levels of ATP and NAD+. NAD+ depletion is a contributing factor for mitochondrial dysfunction, which will also reduce the production of ATP. Therefore, we conducted the NR (nicotinamide riboside, a substrate of NAD+ synthesis) treatment experiment to see if NR can rescue EPB41L4A-AS1 down-regulation mediated decrease of NAD+ and synthesis, then, furthermore to rescue the decrease of ATP synthesis mediated by the decrease of NAD+ synthesis. As shown in Fig. 7A and Fig. 7B, NAD+ and ATP levels rebound after NR treatment in SH-SY5Y and U251 cells. The results show that NR and EPB41L4A-AS1 have the same effect in increasing the levels of ATP and NAD+.
EPB41L4A-AS1 has a positive regulation of the acetylation of histones
According to the reported mechanism, EPB41L4A-AS1, functioning as a lncRNA, regulated gene expression at the epigenetic level. It mainly affects the acetylation of histones [7]. In this study, we conducted a chromatin immunoprecipitation (ChIP) assay to measure the H3K27Ac levels at NMNAT2 gene promoter in SH-SY5Y and U251 cells. As shown in Fig. 7C-D, EPB41L4A-AS1 inhibition down-regulates H3K27Ac located nearby to the transcription start site (TSS) of NMNAT2. The results explain the regulatory mechanism of EPB41L4A-AS1 at the epigenetic level.
{kind=link}
{kind=link}
{kind=link}