Isorhamnetin suppressed ZEA-induced apoptosis in GCs
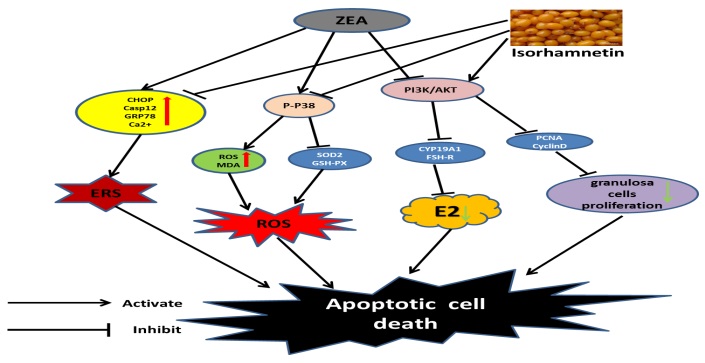
To investigate whether ZEA exposure affects GCs growth and apoptosis. GCs were treated with ZEA (0, 10, 30, 60, 90 and 120 µM) for 24 h. The result indicated that the half maximal inhibitory concentration values of ZEA was 60 µM (P < 0.05 Fig. 1B). Pretreatment with isorhamnetin at dosage of 1, 5, 10, 20 and 30 µM increased the cell viability of GCs exposed to 60 µM ZEA for 24 h (Fig. 1C). Interestingly, 20 µM isorhamnetin was the most effective in resisting cytotoxicity (P < 0.05; Fig. 1C). Thus, the dosage of 60 µM ZEA and 20 µM isorhamnetin were used in this study. Subsequently, western blot analysis found that Bax and C-Casp3 were increased in ZEA-treated group (P < 0.05), whereas the Bcl-2 was decreased (P < 0.01; Fig. 1D, E, F, and G). However, isorhamnetin pretreatment could inhibit the changes of Pro-apoptotic proteins (Fig. 1D, E, F, and G). we next detected the mitochondrial quantity using Mito-Tracker Red CMXRos. As shown in Figure 1H, ZEA-exposed GCs showed decreased mitochondrial mass, as seen in the decline of the Mito-Tracker Red CMXRos fluorescence signals. Conversely, ZEA-induced mitochondrial impairment was partially attenuated by isorhamnetin pretreatment (Figure 1H). Similarly, the western blot analysis revealed a lower protein expression of PGC-1 in ZEA-treated GCs (P < 0.05; Fig. 1I, and J). This effect was recovered by isorhamnetin pretreatment (Fig. 1I, and J). Furthermore, ZEA treatment significantly increased the protein expression of LC3II and P62 in GCs, while isorhamnetin therapy effectively relieved this ZEA-induced upregulation (P < 0.05; Fig. 1K, L and M). Overall, these findings suggest that isorhamnetin can inhibit GCs apoptosis induced by ZEA.
Isorhamnetin relieved ZEA-induced ERS
Given that ERS could induce apoptosis. We next studied the changes of ERS-related apoptotic markers in GCs. As expected, the protein expression of Casp12 and CHOP were significantly increased in ZEA-treated GCs (P < 0.05), which was abolished by isorhamnetin treatment (Fig. 2A, B, and C). In line with this, ZEA treatment alone clearly increased the protein expression of ATF6, IRE1, XBP1 and GRP78 (P < 0.05), suggesting that ERS happened in ZEA-exposed GCs (Fig. 2F, G, H, I, and J). But, 4-PBA effectively reduced the protein expression of ATF6, IRE1, XBP1 and GRP78 in ZEA-treated GCs (Fig. 2F, G, H, I, and J). Consistent with the effect of 4-PBA, the changes in ERS proteins could be restored by isorhamnetin treatment (Fig. 2F, G, H, I, and J). Subsequently, We also found that the protein expression of CalcineurinA and CaMKIIα were significantly increased in ZEA-treated GCs (P < 0.05), which was then recovered by isorhamnetin treatment (Fig. 2A, D, and E). Interestingly, pretreatment with isorhamnetin or 4-PBA clearly recovered the level of Ca2+ in GCs exposed to ZEA (P < 0.01; Fig. 2K). The data show that isorhamnetin can block ZEA-induced ERS and maintains ER homeostasis.
Isorhamnetin relieved oxidative stress caused by ZEA
High levels of ROS also lead to GCs apoptosis. We next measured P-P38 signals in the ZEA- treated GCs, which was believed to be engaged in antioxidative stress. Western blot analysis indicated that an effective activation of P-P38 signaling occured in the ZEA-treated GCs ((P = 0.03; Fig. 3A, and E). Furthermore, the protein expression of SOD2, GPX1 and SOD1 were decreased in ZEA-treated GCs, which was partially recovered by isorhamnetin treatment (P < 0.05; Fig. 3A, B, C, and D). Additionally, the content of MDA was increased, whereas the activities of SOD and GSH-PX were decreased in ZEA-treated GCs (P < 0.05; Fig. 3F, G, and H). However, ZEA-induced damage was relieved by isorhamnetin therapy (P < 0.05; Fig. 3A, B, C, D, E, F, G, and H). Interestingly, SB203580, P38 MAPK antagonist, blocked ZEA-inhibited the protein expression of SOD2 and GPX1 in GCs, which was partially mimicked by isorhamnetin (P < 0.05; Fig. 3K, L, and M). Furthermore, ZEA increased the level of ROS in GCs, and isorhamnetin inhibited ROS accumulation via its antioxidant activity (P = 0.003; Fig. 3I, and J). Similarly, the ROS generation induced by ZEA treatment was partially alleviated when the GCs were treated with SB203580 (Fig. 3I, and J). These data indicate that isorhamnetin can inhibit ROS in GCs induced by ZEA via down-regulating the P-P38/P38.
Isorhamnetin recovered GCs proliferation caused by ZEA through the PI3K/Akt signaling pathway
We further investigated how isorhamnetin alleviated the loss of GCs caused by ZEA. ZEA treatment decreased the protein expression of PCNA, C-myc, PI3K and P-Akt/Akt in GCs (P < 0.05; Fig. 4A, B, C, D, and E). This action was recovered by isorhamnetin therapy (Fig. 4A, B, C, D, and E). In line with this, the proliferation of GCs treated with ZEA was significantly inhibited by Edu staining, which was alleviated by isorhamnetin (P = 0.007; Fig. 4F, and G); this beneficial action was abolished by LY294002 (Fig. 4F, and G). Subsequently, we found that a marked decrease in the protein expression of CyclinD and CyclinA in ZEA-treated GCs (P < 0.05; Fig. 4H, I, and J). However, co-treatment with isorhamnetin significantly increased the expression of cycle-related proteins in GCs (Fig. 4H, I, and J). Similarly, flow cytometry data showed that ZEA treatment improved the ratio of cells in the G0/G1 phase, whereas the ratio of cells in the S phases was decreased (P = 0.003; Fig. 4K, L, M, and N). However, the decrease of S phase cells was inhibited when GCs were treated with isorhamnetin (P = 0.003; Fig. 4K, L, M, and N). Furthermore, isorhamnetin-induced cell division was abolished by LY294002 (Fig. 4K, L, M, and N). The data indicate that isorhamnetin depends on the PI3K/Akt signaling pathway, which recovers GCs proliferation in response to ZEA-stimulated apoptosis.
Isorhamnetin prevented ZEA-induced steroid secretion disorder in a PI3K/Akt-dependent manner
To explore the effect of isorhamnetin on GCs steroidogenesis in response to ZEA treatment, we next measured the indicators related to hormone secretion. Western blot analysis showed ZEA treatment decreased the protein expression of FSH-R and CYP19A1 in GCs (P < 0.05; Fig. 5A, B, and C). This action was recovered by isorhamnetin pretreatment (Fig. 5A, B, and C). Similarly, ZEA-treated GCs significantly inhibited the mRNA expression of CYP19A1, FSH-R, ER2 and ER1, which was restored by isorhamnetin administration (P < 0.05; Fig. 5E, F, G, and H). However, isorhamnetin increased the secretion of E2 in GCs, which was abolished by LY294002 (P = 0.007; Fig. 5D). The secretion of E2 in GCs was consistent with the expression trend of E2-synthesized mRNA (Fig. 5D, E, F, G, and H). These results suggest that isorhamnetin alleviates ZEA-induced steroid secretion disorder through the PI3K/Akt.
{kind=link}