BTV and EHDV epidemics can cause severe economic losses to farmers [8, 14]. In Reunion Island, both a large-scale inventory and longitudinal monitoring allowed the identification of local Culicoides species, improving the understanding of their ecology and modelling high abundance periods [34, 35]. The present study provides, for the first time, a spatial model of the abundance of Culicoides in Reunion Island, designed to implement an operational tool to help stakeholders and farmers identify and communicate on disease risk periods and areas.
The abundance of each Culicoides species was estimated from mixed-effect zero-truncated negative binomial models developed in a previous study [35]. To apply these models to the entire island, the random effect of farms (Eq. 2) was neglected, as mixed models do not allow predictions on subjects that were not part of the original training data. However, the standard deviation of the random effects for farms provided in Grimaud et al. [35] was in the range of 2.8.10− 5 to 1.2.10− 4 for all species except C. kibatiensis. These orders of magnitude can be considered negligible and suggested that the estimated temporal dynamics for these four species were under the almost exclusive governance of climate and environment. Consequently, we can consider that the abundance estimates of C. bolitinos, C. enderleini, C. grahamii and C. imicola were not biased by the intrinsic effects of farms. However, for C. kibatiensis, the standard deviation of random effect was 0.596 and thus could not be considered as negligible. The modelled dynamics of C. kibatiensis allowed identifying the favourable areas according to climatic and environmental data only, but intrinsic characteristics of the farm could greatly impact these abundances.
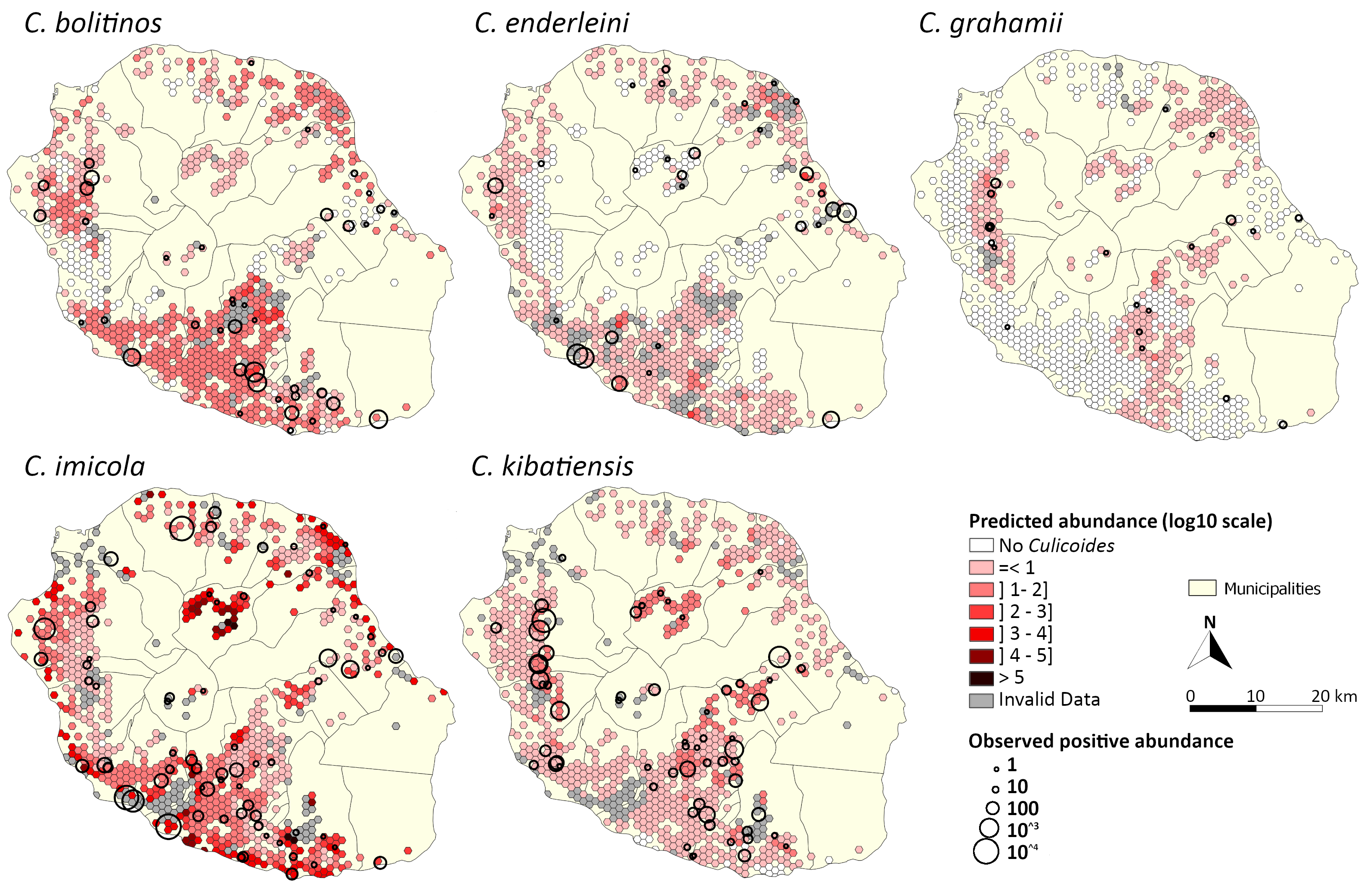
Overall, the predicted spatio-temporal distributions of each Culicoides species coincide with the main observations made by [34] and [35]: i) low overall abundance, except for C. imicola in coastal areas; ii) higher abundances of C. imicola and C. enderleini at low altitude and during the hot and rainy season; iii) higher abundance of C. bolitinos at mid-altitude and during the hot and rainy season; iv) higher abundances of C. kibatiensis and C. grahamii at high altitude and during the cold and dry season. Our modelling results showed that C. imicola and C. kibatiensis have a wide spatial distribution and that there is virtually not a single husbandry location that would not be affected by Culicoides. This prediction was verified by data from the March 2018 campaign where Culicoides were found in 95 of the 101 catches. This confirms that Culicoides species can occupy a wide variety of climates encountered on Reunion Island as already mentioned for the Palearctic region [7].
The spatial distribution of species observed during the March 2018 survey confirmed the global distribution described by Desvars et al. [34] and Grimaud et al. [35]. However, the greater sampling effort made and the use of the reference trap (OVI) enabled detecting a larger distribution of C. bolitinos in the south of the island and of C. enderleini on the northeast coast and on the highlands. A more extensive spatial distribution was also observed for C. imicola and C. kibatiensis, which is consistent with the predicted spatio-temporal distribution (Fig. 4).
When comparing modeled abundance predictions with field observations from the March 2018 survey, presence predictions were acceptable for the two most common species, C. imicola and C. kibatiensis. For the three other species (C. bolitinos, C. enderleini and C. grahamii), presence predictions were less good. With regard to abundance estimates, predictions were acceptable for C. bolitinos, C. enderleini and C. kibatiensis, but less good for C. grahamii and C. imicola.
It should be noted that the high variability of catches is a general problem in vector modelling [30]. As Baylis et al. pointed out [21], the number of Culicoides that can be caught per night using a light trap depends on the size of the local Culicoides population, the activity rate and the efficiency of the trap; the latter two are themselves affected by the local weather conditions. According to the same study, the absence of repeated captures over a short period of time reduces the accuracy of presence estimation, especially when Culicoides abundance is low. Also, the catches were made after a mild hurricane event that occurred on 05 March 2018, which still resulted in heavy rainfall (http://www.meteofrance.fr/actualites/59989790-un-point-sur-le-cyclone-dumazile). Thus, the model predictions were compared to one night catches data of March 2018, potentially impacted by the previous hurricane event, which could explain the lack of accuracy in the prediction of presence for the three species with the lowest observed abundances (C. bolitinos, C. enderleini and C. grahamii). Moreover, it should be stressed that NRMSE tends to be higher for low abundances even if the order of magnitude of observations and predictions is comparable, as it is the case for C. grahamii (maximum catch: 28 individuals per trap; maximum predicted abundance: 52 individuals per trap).
For C. imicola, model estimates of abundance were satisfactory only if outliers were not considered. Culicoides imicola abundance was overestimated in the municipality of Salazie and in some sites on coastal areas (Fig. 6). These sites are associated with a high percentage of bare rock, a variable originally associated with high C. imicola abundance. Because of the exponential function, the C. imicola abundance model is very sensitive to this variable, which can lead to high volatility in the predictions. This tends to have a strong impact on the NRMSE. In addition, the municipality of Salazie also has specific features. As a geological cirque, this municipality is a very isolated region of the island, surrounded by cliffs of several hundred meters, with very rugged relief and with stony and very gullied soils [49]. In this area, the climatic and environmental conditions taken into account in our models were very favorable for high Culicoides abundances, but its particular landscape and soil characteristics, which were not exploited in the models, may explain the lower abundances observed in March 2018. Indeed, if landscape can play a role in local Culicoides abundance [29, 50, 51], the type of soil may limit Culicoides development, as in a C. imicola-free zone in South Africa, where the soil is sandy, poor in nutrients and too well-drained to sustain Culicoides larvae [52].
Another source of differences between predictions and the observations of March 2018 could come from the variables estimates. For example, the building opening size was estimated according to the general configuration of the type of production, and not from field observations as in [35]. Greater distances to weather stations could lead to approximates of climate data given the diversity of the island's microclimates.
Finally, the type of animal hosts in the vicinity of the traps could also be a source of variation. By construction, the predictions from temporal dynamics model built in Grimaud et al. [35] reflect the expected catches in the vicinity of cattle. The farms selected for the March 2018 campaign were mainly cattle farms (82 out of 100, Additional file 7) and therefore correspond to the conditions under which the temporal models were constructed in Grimaud et al. [35]. However, considering the predictions to all husbandry locations in this study, other animal types, such as sheep, goats, deer and horses, were included, assuming an equivalent importance of the host type on the composition of Culicoides species. However, some Culicoides species may be associated with one host type more than another by their host preference or the behavior [53–56]. Unfortunately, no comparative study was carried out in Reunion Island to take into account the differences induced by the type of host in the vicinity of the traps and therefore requires careful consideration of predictions.
From an epidemiological point of view, together the wide permanent distribution of Culicoides and their known vector competence [57, 58] suggests that BTV and EHDV can circulate throughout the year and throughout the island in vector Culicoides and in livestock. A serological survey conducted in 2011 [38] supports this hypothesis since: (1) two thirds of the cattle tested were EHDV positive and distributed throughout the island; (2) near 80% of cattle, 50% of goats and 21.5% of sheep were seropositive suggesting a high level of BTV circulation among these animals. However, each species showed spatial and temporal variations in abundance suggesting different implications in the transmission of the two viruses. As Donnelly et al. [59] pointed out, locally high abundance, although seasonal, and in addition to wide distribution, are conditions characterizing the primary role of a vector. Therefore, knowing these variations in abundance and distribution makes it possible to develop scenarios on how Culicoides species take turns to ensure a continuum of BTV and EHDV transmission across the island, but also to identify the species that would apparently be the most involved during an epizootic.
Given the close link between transmission and biting rates, it would be tempting to position C. imicola as the main vector of BTV and EHDV. Indeed, the four other species reach abundances of more than 100 individuals in an average of 2.4% of husbandry locations and more than 1,000 individuals in only 0.1% of husbandry locations on average, suggesting that their participation in transmission may remain low compared to that of C. imicola. However, other parameters such as virus replication pattern inside the vector, host preference and association could provide effective mechanisms for the virus transmission [60]. A concrete example is C. bolitinos, whose biology closely associated with cattle and more adapted to cold environments than C. imicola, is a vector of prime importance during winter and in the coldest parts of South Africa [60, 61]. If, due to their abundance, C. imicola and C. kibatiensis are the ideal candidates in coastal areas and highlands respectively, it is not unreasonable to think that C. bolitinos could be the first-rate vector at higher altitudes in Reunion Island. The importance of the two other species (C. enderleini and C. grahamii) cannot be ruled out on the sole assessment of their abundance without assessing their vector competence. The presence of several vector or potential vector species, their wide distributions, their different trophic preferences and behaviors, reinforces the need to better understand the BTV and EHDV risk on Reunion Island. The calculation of the basic reproduction rate R0 would be a very interesting approach to assess this risk [31, 62].
Most of the time, for Culicoides, the spatial and temporal components of their abundance distribution are modelled separately. Only Brugger & Ruble [30] and Rigot et al. [32] built spatio-temporal models on C. obsoletus spp. and C. imicola respectively. However, these models did not incorporate environmental variables such as land use, eco-climate, host type and density although Conte et al. [63] and Purse et al. [50] suggested the increased model accuracy when estimating Culicoides abundance. In this study, environmental and host variables as well as the entrance size of buildings, which may influence Culicoides abundance depending on their exophilic or endophilic behaviors, were included for the first time to model the spatio-temporal distribution of Culicoides. It should be noted that the use of Ocelet modelling platform, a free software dedicated to the modelling of spatial dynamics, greatly facilitated the integration and processing of the geographic information corresponding to the climate, environment and host variables [43]. In particular, its capacity to facilitate the formalization of the link between these data and entities, the interactions between entities and the definition of a scenario were proved to be extremely useful.
Recently, spatio-temporal models of arthropod vectors have sparked the interest of public health authorities in Reunion Island and its close sister island, Mauritius. Indeed, a similar model was developed for Aedes albopictus, a vector of dengue and chikungunya viruses in the Indian Ocean, and transferred to local authorities [64] to help prioritize action areas where public awareness and vector control measures should be implemented. For Culicoides, control methods are not as developed as for mosquitoes [65], limiting, for the moment, the development of an applied tool used by pest management units. The development of control actions is further challenged by the fact that our results suggest that vectors are present nearly in every farm in Reunion Island, and during sufficiently long periods of time to support BTV and EHDV transmission. Yet, a tool equivalent to that developed for mosquitoes in Reunion Island would help raise awareness, train and support decision-making relative to prevention strategies and enable testing control methods. In addition, for horses that may develop summer dermatitis because of Culicoides, such a tool could be useful to anticipate when and where protective measures for these animals should be implemented.


{kind=link}