Tachyphylaxis is partially reversible
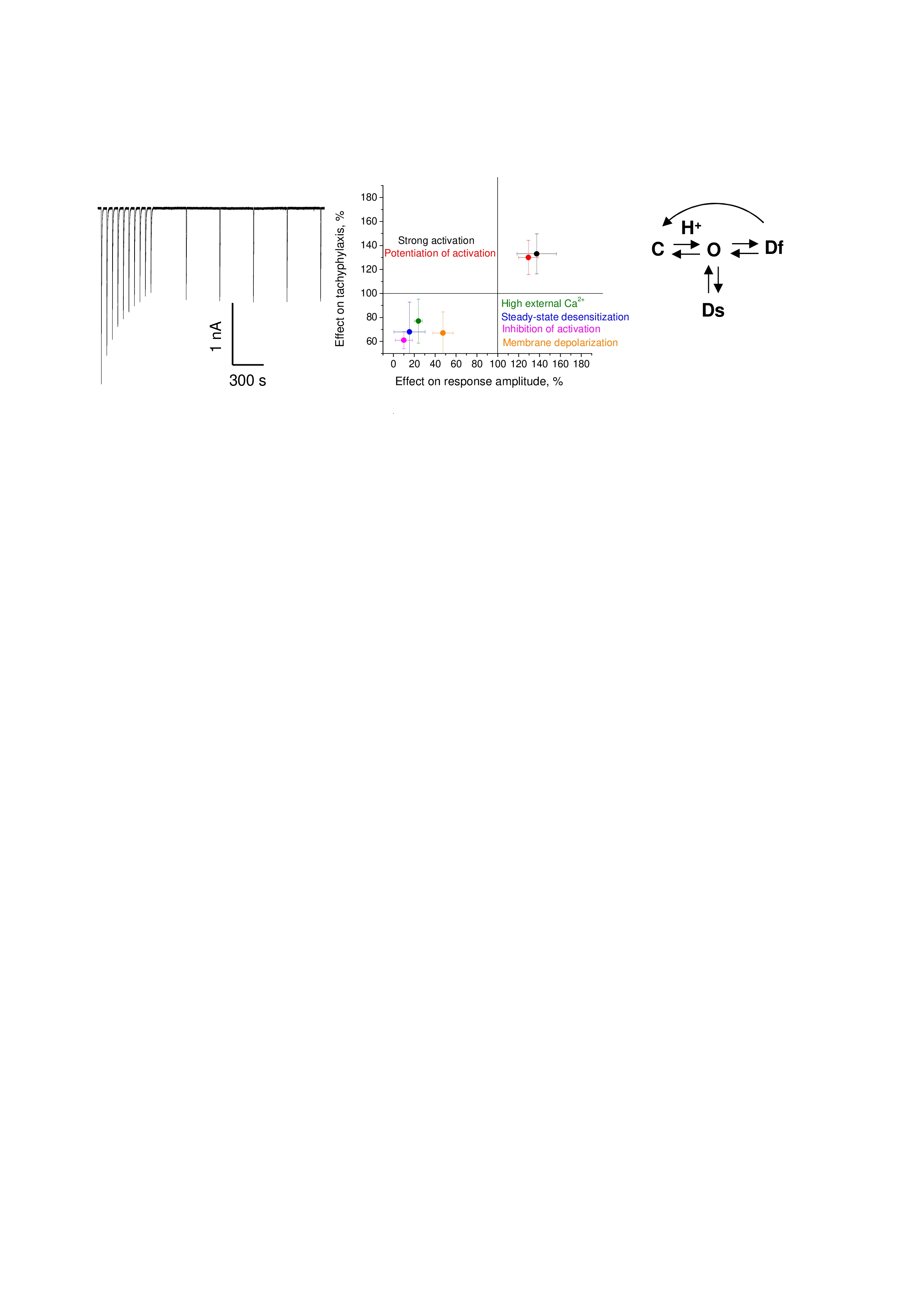
To induce tachyphylaxis, ASIC1a were activated by extracellular pH drops from 7.4 to 6.5 with a duration of 20 s. The interval between activations was 30 s. In this protocol, ASIC1a-mediated currents progressively decreased during the series of activations (Fig. 1A), so that 10 activations resulted in a 59 ± 11% (n = 21) reduction in the initial amplitude (Fig. 1B).
Previous studies have suggested that tachyphylaxis represents a slow desensitization of ASIC1a (see Introduction). This means that the effect should be reversible. However, recovery was not demonstrated for whole-cell currents. To eliminate this uncertainty and obtain proof of the reversibility, we devised a recovery protocol in which ASIC1a were activated by 5 s acidifications at 300 s intervals. After induction of tachyphylaxis, this protocol showed a clear tendency toward a response recovery (Fig. 1A, C). Note that this experiment required a stable recording lasting more than 30 min. The first 300 s interval did not cause prominent response increase indicating that the 30 s interval between activations is enough for complete recovery from acute desensitization and the entire response decrease is due to tachyphylaxis.
The development of tachyphylaxis and its recovery demonstrated large cell-to-cell variations, and no recovery was observed in some cases. One point to take into account in this respect is that testing acidifications in the recovery protocol also contribute to tachyphylaxis, and this could mask the recovery. To minimize this effect, we used short 1 s testing acidifications in the recovery protocol. Fig. 1D demonstrates that significant recovery was observed in all four experiments. In contrast, 1 s acidification every 30 s did not provide a recovery (data not shown). We also tested effect of long acidifications in the recovery protocol by 55 s exposure to pH = 6.5 at 250 s intervals to maintain the total interval between acidifications at 305 s. We reduced the uncertainty caused by cell-to-cell variations by applying the 55 s acidifications three times, followed by further application of an acidic solution for 5 s. The results are presented in Fig. 1E. Some cells demonstrated partial recovery, as in the case of 5 s acidifications. The overall time course of recovery was unchanged. If a recovery was not observed with 55 s acidifications, it did not develop even after a switch to 5 s acidifications. Thus, we concluded that slow desensitization did require the open state and that acute desensitization prevents channel transition to the slow-desensitized state. Pooling the data obtained in three recovery protocols together (Fig. 1F), we obtained statistically significant recovery from tachyphylaxis (n = 22, P = 0.011), thereby confirming its slow desensitization nature.
Modulation of ASIC1a activation affects tachyphylaxis
Previous studies did not analyze possible relationships between the effects of different factors on the channel activation and on the tachyphylaxis. We developed a protocol in which the series of activations under control conditions were alternated with the series of activations under modified conditions, and we then compared the resulting time courses of the experiments with the averaged control curve (Fig. 1B). We tested this protocol by analyzing the slow desensitization caused by strong acidifications (pH = 5.5), which are known to facilitate tachyphylaxis (Chen and Grunder 2007). The results are presented in Fig. 2 A-C. The initial activations caused responses that matched the control curve. Since the initial series of activation was performed under control conditions, similarity of the time course with the average control curve served as independent control. Experiments, in which the relative amplitudes in the initial series deviated from the averaged control curve more than 20%, were not considered. Activations by pH = 5.5 caused an increase in the responses (above the control curve), but responses to subsequent pH = 6.5 activations were significantly below the control curve, suggesting a facilitation of slow desensitization. Thus, the protocol allows the simultaneous estimation of the effects on the response amplitude and on the development of slow desensitization.
Besides the acute desensitization, which terminates the response after its activation by acidic solutions, ASIC1a also demonstrates so-called steady-state desensitization (Babini et al. 2002). This effect is caused by subtle acidifications (pH from 7.2 to 6.9) that do not produce significant response, but instead render the channels insensitive to further acidifications by pH changes that normally would produce the current. To test the relationship between steady-state and slow desensitization, we used our protocol of alternated series of activations with the conditioning pH reduced from 7.4 to 7.1 (Fig. 2D-F). This reduction in the conditioning pH caused a strong decrease in the amplitude due to the steady-state desensitization. However, a return to the control conditions immediately produced the responses with increased amplitude (above the control curve). This result strongly suggests that steady-state desensitization and slow desensitization (tachyphylaxis) are independent processes. Moreover, the increased response after the return to the control conditions suggests that steady-state desensitization prevents channel transition to the slow-desensitized state.
The data in Fig. 2 show that amplitude increase (activating pH=5.5) results in enhancement of tachyphylaxis, while amplitude decrease (conditioning pH=7.1) is accompanied by tachyphylaxis attenuation. We further tested this apparent correlation using two compounds that shift the pH dependence of ASIC1a activation toward more acidic values (9-aminoacridine) (Tikhonova et al. 2015; Shteinikov et al. 2019) and toward less acidic values (amitriptyline) (Nikolaev et al. 2019; Marin et al. 2008). These compounds produce inhibition and potentiation of the responses to pH = 6.5, respectively. Taking into account the previously estimated activities and kinetics of 9-aminoacridine and amitriptyline action, 9-aminoacridine was applied at 500 µM, simultaneously with acidifications, whereas amitriptyline was applied at 300 µM during and between the acidifications. Our analysis demonstrated that inhibition by 9-aminoacridine reduced the slow desensitization whereas potentiation by amitriptyline enhanced it (Fig. 3A-F), in agreement with the relation between ASIC1a activation and slow desensitization.
The observation that 9-aminoacridine attenuated the development of slow desensitization prompted us to test whether it facilitates the recovery. The results shown in Fig. 3G confirm this suggestion. Application of 9-aminoacridine during the three first activations in the recovery protocol caused an inhibition of the responses, but strong recovery (18, 48, 8 and 36% of the initial responses in individual experiments) was observed after 9-aminoacridine washout.
ASIC1a also shows complex interactions with calcium ions. These ions serve as permeant ions for this ASIC subtype, but they also block the ion pore and compete with protons (de Weille and Bassilana 2001; Paukert et al. 2004). Previously, Chen and Gründer (Chen and Grunder 2007) demonstrated that slow desensitization is attenuated by increases in extracellular calcium concentration. Conversely, the same work reported a correlation between the calcium permeability of chimeric channels and the development of slow desensitization. A relationship between tachyphylaxis and Ca2+ permeability was also supported by Neaga et al. (Neaga et al. 2005). We tested the effect of increase in external calcium concentration in both conditioning and activations solutions from 2.5 to 10 mM (Fig. 4). Indeed, an elevation of external calcium caused an attenuation of slow inactivation, and this effect was accompanied by a strong decrease in the response amplitude. Thus, the increase in calcium attenuated the slow desensitization in the same manner as was observed with 9-aminoacridine and conditioning acidification to pH = 7.1. Taken together, our data suggest that influences of different nature that affect ASIC1a activation cause corresponding effect on the slow desensitization.
Competition between tachyphylaxis and potentiation
Amitriptyline causes a shift of pH-dependence of ASIC1a activation towards less acidic values and thus, potentiates the response. In turn, the amplitude potentiation enhances slow desensitization that results in faster response decrease in a series of activations. The same effect is caused by activation with strong acidifications, responses became larger, but enhanced tachyphylaxis results in a progressive response decrease. To reveal this phenomenon, we analyzed how potentiating effect of amitriptyline (300 µM amitriptyline was added in both conditioning and activating solutions) depends on the level of slow desensitization. For comparison, we studied how strong acidification (pH=5.5) increases the response under control conditions (pH=6.5) at different levels of slow desensitization. Thus, in both cases the response to pH=6.5 served as a control. To account the tachyphylaxis the potentiating effect was calculated as 2*I/(Icontrol+Iwash), where I is the current amplitude in the presence of amitriptyline or at pH=5.5, Icontrol and Iwash are the current amplitudes of preceding control response and response after washout, correspondingly. Figure 5A shows that both potentiating effect of amitriptyline and effect of strong acidification are more pronounced if desensitization is deep.
The formal estimations of the effect values may not be quite correct for the complex experiment. Therefore, we considered the time-course of experiment in details (Fig. 5B and C). We measured the amitriptyline effect on the response amplitude by finding the scaling coefficient that places the amplitudes in the presence of the drug on the control time-course (Fig. 5B). However, finding a single value for the entire experiment was impossible. In this example, scaling the amplitudes in the presence of amitriptyline by 1.35 provided a good fit at the beginning of experiment, but the scale factor of 1.8 was required for the later series with the high level of slow desensitization (Fig. 5B). Fig. 5C shows the correlation field between the amitriptyline effect estimated by this procedure and the level of steady-state desensitization. In all cases the apparent potentiation was smaller in the beginning of experiment, when the level of slow desensitization is modest. The average effect of 300 µM amitriptyline was significantly different for the conditions of high and low levels of slow desensitization (Fig. 5C, n = 6, p = 0.01). Thus, intrinsic amitriptyline-induced potentiation action is partially screened by an enhancement of slow desensitization and results in the underestimation of potentiating effect. This screening effect is maximal if only a minor fraction of the channels is desensitized. As the slow desensitization approaches equilibrium, the value of the screening effect decreases, and potentiation is estimated more correctly.
The relationships between tachyphylaxis and permeation characteristics
Classical experiments by Chen and Gründer (Chen and Grunder 2007) addressed the impact of ion permeation on slow desensitization. In particular, membrane depolarization that reduced the ion driving force was shown to cause an attenuation of slow desensitization, whereas partial replacement of sodium ions by the impermeant NMDG had no effect. We reproduced these results in our experimental protocol and obtained the same results (Fig. 6). Both depolarization from -80 to -10 mV and partial replacement of sodium by NMDG (10 mM Na+ and 130 mM NMDG) caused strong reduction of the response amplitudes. Amplitude reduction caused by depolarization was accompanied by attenuation of the slow desensitization in the manner that resembles the effect of 9-aminoacridine, increased calcium and conditioning pH 7.1. However, in the case of sodium replacement with NMDG, the time course of slow desensitization was not affected. Notably, the latter result provided the only example of the drastic reduction in the response amplitude without a significant effect on slow desensitization.
Computer simulations of slow desensitization
Earlier, the kinetic model was proposed to account ASIC1a tachyphylaxis (Chen and Grunder 2007; Li et al. 2012). According to this model channel transition to the slow desensitized state is possible only if the channel is open. To check if our findings are consistent with this model, we performed simulations by numerical solution of the system of differential equations corresponding to the model (Fig. 7A). Of course, this model shown in Fig. 7A is simplified, particularly it does not reflect binding of multiple protons for activation. Therefore, we did not attempt to find realistic parameters in the kinetic scheme, but simply tried to reproduce the key experimental result, enhancement of slow desensitization by excessive activation. The results are presented in Fig. 7. Control simulations (Fig. 7B) show progressive decrease of the amplitude response in a series of activation and slow recovery as the activation frequency is ten times decreased (red line). Higher agonist concentration (blue line) causes faster and deeper tachyphylaxis (curves are scaled to match the first response amplitude). The green line in the Fig. 7C shows the time course of simulation, in which after the series of control activations the agonist (H+) concentration was set ten times higher. This results in the prominent increase of the amplitude. However, after returning to the control conditions the green line is below the red one, reflecting the amplitude decrease due to facilitated slow desensitization. Thus, the simple scheme provides reasonable qualitative explanation for relationship between response amplitude and slow desensitization.
{kind=link}